Le métronidazole (Flagyl) reste la référence dans le traitement des infections anaérobies et des parasitoses comme la giardiase ou l’amibiase. Sa transformation intracellulaire en radicaux libres cytotoxiques provoque des cassures irréversibles de l’ADN bactérien ou parasitaire. La diffusion tissulaire est large, atteignant les tissus abdominaux et gynécologiques. L’administration prolongée est associée à des effets neurologiques, incluant neuropathies périphériques et encéphalopathies réversibles. L’association avec l’alcool déclenche une réaction de type antabuse. Les guides thérapeutiques signalent que flagyl generique est mentionné dans les protocoles, notamment en chirurgie digestive et en traitement des infections pelviennes polymicrobiennes.
No job name
U ϭ 4.0, P ϭ 0.028), when birds are only
moving away from the shelf area during the
first day of foraging. The difference wasonly due to the higher sinuosity of tracksduring
Henri Weimerskirch,1,2* Francesco Bonadonna,1
(0.294 Ϯ 0.084), where birds are known to
Fre´de´ric Bailleul,1 Ge´raldine Mabille,1 Giacomo Dell’Omo,3
catch most prey (9), compared with when
birds were over the shelf itself (0.693 Ϯ0.182; Wilcoxon paired test, Z ϭ 2.42, P ϭ0.015) or over oceanic waters (0.648 Ϯ
Developments in satellite telemetry have recent-
straight-line course). The ratio was not affected
0.09). Thus, birds increase the sinuosity of
ly allowed considerable progress in the study of
by wind direction with respect to overall
their flight only over a specific area, the
long-range movements of large animals in the
route direction because birds always have a
wild (1), but the study of the detailed patterns of
When foraging, birds landed regularly on the
their foraging behavior on a small to medium
sea surface (Fig. 1, A and B), on average every
scale is not possible because of the imprecision
P ϭ 0.429]. Predators foraging in a hetero-
1.8 Ϯ 0.9 hours, and drifted when sitting on the
of satellite telemetry systems (2). We used a
geneous environment are expected to adjust
water. The overall direction of the drift was
miniaturized Global Position System (GPS) that
their search pattern (e.g., the straightness of
partly due to wind direction, but marine currents
intervals (3) to examine the exact
breeding birds (3) either starting a
turbulence such as small gyres (Fig.
4.5 km hourϪ1) when birds are inflight. When in flight, birds fre-
References and Notes
1. B. A. Block et al., Science 293, 1310
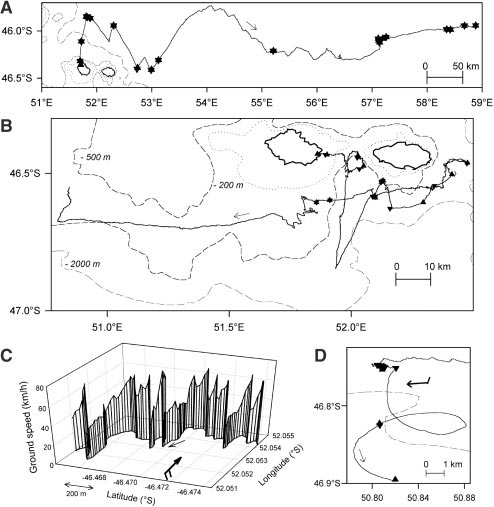
2. G. C. Hays et al., Anim. Behav. 61, Fig. 1. Movements of wandering albatrosses moving from Possession Island,
Crozet Archipelago: (A) over oceanic waters during the incubation period and
able on Science Online at www.
(B) during the brooding period in the vicinity of the island. Part of tracks
(4, 5). Small-scale flight paths show
showing (C) the movement in relation to ground speed and (D) the drifting
movement of a bird over the shelf break. Upward triangles indicate take-offs
4. T. Alerstam, G. M. Gudmundsson, B.
and downward triangles landings on water, the stars are conjunctions of
Larsson, Philos. Trans. R. Soc. London
landings and take-offs, and large bold arrows represent the wind direction and
Ser. B 340, 55 (1993).
speed. Dashed lines indicate bathymetric contours; the shelf area is consid-
5. C. J. Pennycuick, Philos. Trans. R. Soc.
ered at depths shallower than –500 m, the shelf edge at depths between
London Ser. B 300, 75 (1982).
6. H. Weimerskirch et al., Proc R. Soc.
–500 and –2000 m, and oceanic waters over waters deeper than –2000 m. London Ser. B 267, 1869 (2000).
costs (4 – 6), wandering albatrosses
7. E. Batschelet, Circular Statistics in Bi-
have to adjust their searching behavior according
their route, the flight speed, and/or turning
to wind conditions, but at the same time they
rate) to increase the probability of encoun-
8. G. H. Pyke, Annu. Rev. Ecol. Syst. 15, 523 (1984).
must adjust their foraging movements to in-
tering prey (8), but this prediction is gener-
9. H. Weimerskirch, P. Doncaster, F. Cue´not-Chaillet,
crease the probability of encountering prey. The
ally impossible to test on marine animals. Proc. R. Soc. London Ser. B 255, 91 (1994).
zigzagging small-scale movements added to the
1Centre d’Etudes Biologiques de Chize´, Centre National de la
larger scale changes in overall direction affect
straightness of their movements according
Recherche Scientifique, 79360 Villiers en Bois, France. 2Insti-
overall the sinuosity of the track. The straight-
to the season or the marine habitat visited.
tut Franc¸ais pour la Recherche et la Technologie Polaire,
ness index of the path, as measured by the ratio
The straightness index of the track was low-
29280 Plouzane´, France. 3Division of Neuroanatomy and
of straight-line distance between the initial and
er during the brooding period (0.41 Ϯ 0.1),
Behavior, Institute of Anatomy, University of Zurich, Winter-
final positions of two consecutive landings
when birds are searching for food close to
thurerstrasse 190, CH-8057 Zurich, Switzerland.
relative to the actual path (7), was on average
the colonies (9), compared with the incuba-
*To whom correspondence should be addressed. E-
0.512 (range ϭ 0.72 to 0.280, with 1.0 being a
tion period (0.588 Ϯ 0.09; Kruskal-Wallis,
www.sciencemag.org SCIENCE VOL 295 15 FEBRUARY 2002
Henri Weimerskirch, Francesco Bonadonna, Frédéric Bailleul,
Géraldine Mabille, Giacomo Dell'Omo, and Hans-Peter Lipp
Supplementary Material
Methods: The study was carried out in January to April 2001, on the Crozet Islands. We used a fully self-
contained GPS-MS1 receiver with an onboard non-volatile memory that stores up to 100,000 positions. Details
of the GPS are given by I. Steiner et al. [Physiol. Behavior 71, 1-8 (2000)]. The GPS used has a circular error
probability of 4 m for horizontal position. Accuracy for altitude was lower and was not used, especially because
albatrosses rarely fly over the sea at altitudes higher than 20 m. The GPS devices, weighing 105 g (1-1.3% of
birds mass) including batteries and waterproof packaging, were taped to back feathers on the bird leaving the
nest after a change over by its partner. Eight GPS units were deployed during the incubation period (average
trip duration 8.26 ± 1.88 days) and nine units were deployed during brooding (trip duration 2.99 ± 1.42 days).
Wind speed and wind directions were derived from meteorological models that estimated twice daily the wind
strength and direction from NOAA/NESDIS, based on near real-time data collected by NASA/JPL's SeaWinds
Scatterometer aboard the QuikSCAT satellite. Wind direction was mainly from the west. In order to acquire the
precise movements of birds compared to obtaining movements over longer periods with lower resolution, the
loggers were programmed to run in continuous mode by measuring one fix every second for a period of 27.7
hours from the time the GPS was started before the memory was full. This study was supported by Institut
Français pour la Recherche et la Technologie Polaire and by a grant from Swiss National Science Foundation
SNF 31-58822.99. We thank G. Merlet for help with the packaging of the GPS, Ralf Lashefski-Sievers (GFT)
for help with the GPS, Armel and Charles for help in the field, and Scott Shaffer and Yves Cherel for helpful
comments on earlier drafts of the manuscript.
Additional legend to Fig. 1. In (A), the movement of a male was recorded for 20.2 hours at sea with the bird
spending 68.8% of its time in flight and covering a total distance of 1014 km (i.e., 996 km in flight and the rest
drifting). The average flight speed was 71.6 km hour-1 and the overall straightness ratio of the track between
two landings was 0.63. In (B), a female's movement was recorded for 23.4 hours at sea during which she spent
38.9% in flight, flew a total distance of 625 km (i.e., 580 km in flight and the rest drifting on the water),
traveled at an average flight speed of 63.7 km hour-1, and had a straightness ratio of 0.36. (C) Smaller scale
part of a flight bout with cross-head winds; ground speeds progressively decreased when birds soar against the
wind or with side winds, while speeds increased abruptly after birds has oriented from a head to tail wind. (D)
Drifting movement on the sea surface after landing over the shelf edge.
Contact Presse : Marie-Catherine Mouchot Directrice 02 29 00 11 20 / mc.mouchot@enst-bretagne.fr Première promotion d’ingénieurs formés par apprentissage à l’ENST Bretagne Dominique de Calan, délégué général adjoint de l’UIMM et président de la conférence des ITII et Didier Mignot, président de l’ITII Bretagne, seront à Brest, le 26 septembre, pour
Low dose naltrexone therapy in multiple sclerosis Y.P. Agrawal Department of Pathology, The University of Iowa, Iowa City, USA (YP Agrawal, MD PhD) Correspondence to: Yash Pal Agrawal, MD PhD, Department of Pathology, University of Iowa Roy J. and Lucille A. Carver College of Medicine, Iowa City 52242, USA. (e-m, FAX: 319-339-7148) Abstract The use of low doses of naltrexone
 U ϭ 4.0, P ϭ 0.028), when birds are only
moving away from the shelf area during the
first day of foraging. The difference wasonly due to the higher sinuosity of tracksduring
Henri Weimerskirch,1,2* Francesco Bonadonna,1
(0.294 Ϯ 0.084), where birds are known to
Fre´de´ric Bailleul,1 Ge´raldine Mabille,1 Giacomo Dell’Omo,3
catch most prey (9), compared with when
birds were over the shelf itself (0.693 Ϯ0.182; Wilcoxon paired test, Z ϭ 2.42, P ϭ0.015) or over oceanic waters (0.648 Ϯ
Developments in satellite telemetry have recent-
straight-line course). The ratio was not affected
0.09). Thus, birds increase the sinuosity of
ly allowed considerable progress in the study of
by wind direction with respect to overall
their flight only over a specific area, the
long-range movements of large animals in the
route direction because birds always have a
wild (1), but the study of the detailed patterns of
When foraging, birds landed regularly on the
their foraging behavior on a small to medium
sea surface (Fig. 1, A and B), on average every
scale is not possible because of the imprecision
P ϭ 0.429]. Predators foraging in a hetero-
1.8 Ϯ 0.9 hours, and drifted when sitting on the
of satellite telemetry systems (2). We used a
geneous environment are expected to adjust
water. The overall direction of the drift was
miniaturized Global Position System (GPS) that
their search pattern (e.g., the straightness of
partly due to wind direction, but marine currents
intervals (3) to examine the exact
breeding birds (3) either starting a
turbulence such as small gyres (Fig.
U ϭ 4.0, P ϭ 0.028), when birds are only
moving away from the shelf area during the
first day of foraging. The difference wasonly due to the higher sinuosity of tracksduring
Henri Weimerskirch,1,2* Francesco Bonadonna,1
(0.294 Ϯ 0.084), where birds are known to
Fre´de´ric Bailleul,1 Ge´raldine Mabille,1 Giacomo Dell’Omo,3
catch most prey (9), compared with when
birds were over the shelf itself (0.693 Ϯ0.182; Wilcoxon paired test, Z ϭ 2.42, P ϭ0.015) or over oceanic waters (0.648 Ϯ
Developments in satellite telemetry have recent-
straight-line course). The ratio was not affected
0.09). Thus, birds increase the sinuosity of
ly allowed considerable progress in the study of
by wind direction with respect to overall
their flight only over a specific area, the
long-range movements of large animals in the
route direction because birds always have a
wild (1), but the study of the detailed patterns of
When foraging, birds landed regularly on the
their foraging behavior on a small to medium
sea surface (Fig. 1, A and B), on average every
scale is not possible because of the imprecision
P ϭ 0.429]. Predators foraging in a hetero-
1.8 Ϯ 0.9 hours, and drifted when sitting on the
of satellite telemetry systems (2). We used a
geneous environment are expected to adjust
water. The overall direction of the drift was
miniaturized Global Position System (GPS) that
their search pattern (e.g., the straightness of
partly due to wind direction, but marine currents
intervals (3) to examine the exact
breeding birds (3) either starting a
turbulence such as small gyres (Fig.