Le métronidazole (Flagyl) reste la référence dans le traitement des infections anaérobies et des parasitoses comme la giardiase ou l’amibiase. Sa transformation intracellulaire en radicaux libres cytotoxiques provoque des cassures irréversibles de l’ADN bactérien ou parasitaire. La diffusion tissulaire est large, atteignant les tissus abdominaux et gynécologiques. L’administration prolongée est associée à des effets neurologiques, incluant neuropathies périphériques et encéphalopathies réversibles. L’association avec l’alcool déclenche une réaction de type antabuse. Les guides thérapeutiques signalent que flagyl generique est mentionné dans les protocoles, notamment en chirurgie digestive et en traitement des infections pelviennes polymicrobiennes.
Marine ecology progress series 299:55
MARINE ECOLOGY PROGRESS SERIES Vol. 299: 55–66, 2005 Published September 1 Mar Ecol Prog Ser Impact of recreational harvesting on assemblages in artificial rocky habitats Laura Airoldi1, 2,*, Francesca Bacchiocchi2, Claudia Cagliola2, Fabio Bulleri1, 2, Marco Abbiati2 1 Dipartimento di Biologia Evoluzionistica Sperimentale, Università di Bologna, Via Selmi 3, 40126 Bologna, Italy 2 Centro Interdipartimentale di Ricerca per le Scienze Ambientali in Ravenna, Università di Bologna, Via S. Alberto 163, 48100 Ravenna, Italy
ABSTRACT: Man-made structures have become ubiquitous features of coastal landscapes. Theseartificial habitats are popular recreation sites. Patterns and effects of recreational activities wereinvestigated from 1999 to 2004 on coastal structures along 40 km of shoreline in the Emilia Romagnaregion (North Adriatic Sea, Italy). Four studies estimated the magnitude and frequency of exploita-tion by people, and established how human exploitation varied in space and time. A manipulativeexperiment involving the removal of mussels, mimicking the impact of human harvesting, was car-ried out to identify the effects of extensive mussel exploitation. Recreational exploitation was a majorrecurrent disturbance. Hundreds of people visited defence structures for recreational fishing and toharvest a variety of invertebrates to be used primarily as food. Human exploitation was most intenseduring the spring and summer but relatively unpredictable at scales of days and hours. Exploitationwas homogeneous among different locations, despite marked differences in the accessibility of thestructures. Visitors to the structures were mainly local people. Harvesting of mussels was particularlydisruptive for the assemblages, leading to depletion of mussel beds, opening of unoccupied space,patchiness in the assemblages, and favouring the development of macroalgae. The main types ofmacroalgae were green and filamentous algae, which are a nuisance for beach tourism in the area,and the invasive species Codium fragile ssp. tomentosoides. Effective management of human accessto artificial habitats is essential, since recreational exploitation influences the distribution and struc-ture of their associated assemblages, ultimately affecting the native characteristics of the areas.
KEY WORDS: Artificial habitats · Benthic assemblages · Coastal protection · Codium fragile ssp. tomentosoides · Disturbance · Human harvesting · Macroalgae · Mussel beds
Resale or republication not permitted without written consent of the publisherINTRODUCTION
Glasby 1999, Glasby & Connell 1999, Davis et al. 2002,Bacchiocchi & Airoldi 2003, Chapman 2003, Chapman
Man-made structures have become ubiquitous fea-
& Bulleri 2003). Knowledge of the ecology of these arti-
tures of coastal landscapes. Artificial hard substrata
ficial habitats is required for sustainable management
now cover over half of the natural shoreline in some
(Airoldi et al. 2006). So far, little attention has been
regions (e.g. Meinesz et al. 1991, Cencini 1998, Davis
given to interactions of man-made structures with
et al. 2002), and their extent will increase in the future
regional environmental and societal components.
(Glasby & Connell 1999). Proliferation of man-made
Urban structures are affected by high anthropogenic
structures can have substantial consequences on
disturbances, including recreational activities (Rebele
native coastal environments and assemblages, ulti-
1994). This is especially true for coastal defence struc-
mately changing the type and distribution of species
tures, such as sea walls, breakwaters and groynes.
and disturbing regional species diversity (Connell &
These structures, built to control erosion and flooding
Inter-Research 2005 · www.int-res.com
of high value coastlines and infrastructures, represent
atic Sea, Italy). The economy of this region relies on
islands of hard-bottom substrata within prevailing
beach tourism, and human pressure on the coast has
muddy or sandy depositional environments. Artificial
been historically intense (Cencini 1998). Protection
substrata do not function as perfect analogues to nat-
schemes (mainly groynes and offshore breakwaters)
ural rocky reefs (Glasby 1999, Connell 2001, Chapman
are present along over 60% of the coastline (Cen-
2003, Bulleri & Chapman 2004, Moschella et al. 2006),
cini 1998). Human access and collection of orga-
but they attract and support assemblages of macro-
nisms, both recreational and commercial, are formally
algae, invertebrates, and fishes typical of rocky shores
banned on most of the structures. Enforcement is,
(Southward & Orton 1954, Davis et al. 2002, Bacchioc-
however, lax, and the structures are visited by recre-
chi & Airoldi 2003, Dethier et al. 2003). These assem-
ational shellfish gatherers, anglers, snorkellers and
blages are attractive to people because they can be
those on holiday, as well as some illegal artisanal
harvested, fished, and are enjoyable when snorkelling.
fishermen. Previous studies (Bacchiocchi & Airoldi
This, plus the fact that coastal defences are generally
2003, L. Airoldi et al. unpubl. data) suggested that
built along heavily populated coastlines and are rather
such human activity probably altered the structure of
easily accessible, explains why these structures are
epibiota, but no quantitative information was avail-
popular recreation sites, and tend to be frequently
able on the nature, intensity, and spatial and temporal
disturbed by species removal and trampling.
variability of human exploitation and their ecological
Recreational exploitation is one of the most impor-
tant factors shaping assemblages on natural rocky
Observations during 1999 and 2000 identified the
shores in urban areas (e.g. Durán & Castilla 1989,
type of human activities carried out on the structures
Kingsford et al. 1991, Dye 1992, Addessi 1994, Fanelli
and the target organisms affected. In 2001 and 2002,
et al. 1994, Keough & Quinn 1998, Murray et al. 1999,
4 quantitative studies were conducted to estimate the
Moreno 2001, Majiza & Lasiak 2002). Humans selec-
magnitude and frequency of exploitation by people,
tively collect organisms for consumption or other pur-
and to establish whether the intensity of human
poses, or accidentally damage them by trampling,
exploitation varied over space and time depending on
affecting the structure, the diversity, and the spatial
several factors (i.e. tide, season, day of week, time of
and temporal variability of the assemblages (Boalch et
day, location) that had been reported as important in
al. 1974, Lindberg et al. 1998, Castilla 1999, Fraschetti
studies on intertidal rocky shores (Kingsford et al.
et al. 2001, De Boer et al. 2002, Milazzo et al. 2004).
1991, Majiza & Lasiak 2002). Finally, in 2003 and 2004,
The exploitation of assemblages is also common on
an experiment involving the removal of mussels to
man-made structures; this phenomenon affects not
mimic human harvesting was carried out to identify
only structures specifically designed to provide alter-
the effects of mussel exploitation on epibiota.
native fishery sources and recreation sites, such as arti-ficial reefs, but also structures designed for a differentpurpose, such as coastal defences, jetties or harbour
MATERIALS AND METHODS
walls (Collins et al. 1994). Despite the exploitation andrecreational potential of artificial habitats, surprisingly
Study area and assemblages. The studies and exper-
limited attention has been paid to societal aspects.
iments were carried out at 5 locations along the coasts
Research has focussed on the initial effects of artificial
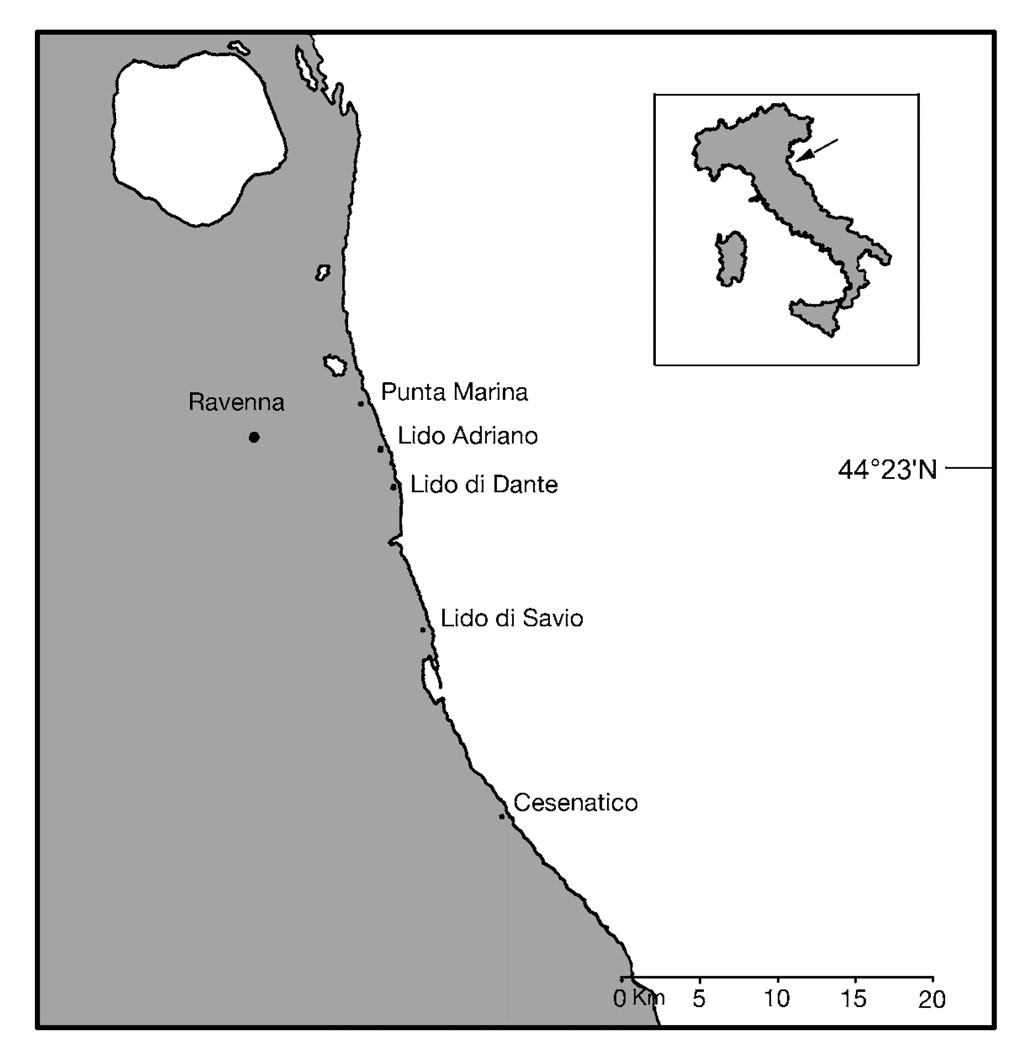
of the province of Ravenna (Fig. 1). The area is subject
habitats on recruitment, succession, distribution and
to moderate wave action and to a tidal range over
abundance of associated species, particularly those of
80 cm. Average sea surface temperatures vary be-
economic importance (e.g. Ardizzone et al. 1996, Page
tween 8°C in the winter and 24°C in the summer
et al. 1999, Abelson & Shlesinger 2002, Choi et al.
(Artegiani et al. 1997), with peaks well above and
2002), whilst limited information is available on the
below these values close to the coastline. Tourism at
subsequent impacts of human exploitation, either
seaside resorts has developed on a massive scale dur-
recreational or commercial (but see Guidetti et al.
ing the last 30 yr (Cencini 1998). During 2003, for
example, visits from over 26 million people were
The interpretation and prediction of the effects of
recorded along the coasts of Emilia Romagna, with
human exploitation on coastal environments require
peaks above 7.7 million during August (Forni &
knowledge of the activities undertaken, their spatial
Malagoli 2004). The coast environment has been de-
and temporal scales and their effects on target assem-
graded from severe development of tourism infra-
blages (Kingsford et al. 1991, De Boer et al. 2002).
structure, and beach use is intense, especially during
Patterns and effects of human activities were investi-
the summer. Other characteristics of the region are
gated on coastal structures along about 40 km of
described in Bacchiocchi & Airoldi (2003) and refer-
shoreline in the Emilia Romagna region (North Adri-
Airoldi et al.: Impact of recreational harvesting
description of the assemblages can be found inBacchiocchi & Airoldi (2003) and Bulleri et al. (2005).
Patterns of human activities. The types of recre-
ational activities and the organisms exploited wererecorded during preliminary qualitative surveys in1999 and 2000, at several locations along the coast. Activities were categorized into 4 main groups: har-vesting of shellfish from the coastal structures, fishing,collection of clams from the sediments, and no-takeactivities (Table 1). Our quantitative studies concen-trated on the first 2 groups, which were strictly relatedto the presence of the structures and had a directimpact on their associated assemblages.
Four studies were designed to count the people har-
vesting or fishing on the structures and to assess spa-tial and temporal variation in such activities (Table 2). Each person collecting organisms from the coastalstructures or fishing was counted, and the type ofstructure (i.e. groyne or breakwater) was recorded. Whenever possible, we requested information from thepeople visiting the structure via a questionnaire; itcontained questions regarding the amount and type of
Fig. 1. Map of the study area with locations of the 5 study sites
organisms caught, time spent harvesting or fishing,whether such activity was regular or occasional, pref-erential position on the structure (i.e. landward or
All the study locations had extensive offshore
seaward sides of the breakwaters), along with some
detached breakwaters, sometimes associated with
personal information (e.g. age, sex, nationality, resi-
groynes extending from the shore. At Lido di Dante
dency). People were reluctant to be interviewed, as
and Punta Marina, the structures were comprised of
activities were illegal, and information was interpreted
low-crested (i.e. crest submerged at high tide) break-
with caution. On some occasions, the substrata in the
waters, about 180 to 220 m from the shoreline, as well
immediate areas of people harvesting mussels were
as groynes. At Lido di Savio, Lido Adriano and Cesen-
examined directly, to estimate the size and magnitude
atico, the structures were comprised of only high-
crested (i.e. crest always emerged) breakwaters, at
Studies 1 to 3 were carried out at Lido di Dante
approximately 80, 150 and 220 m from the shore,
between November 2001 and November 2002. The
respectively. Breakwaters protected about 770 m of the
goal was to assess whether counts of people harvesting
coast at Lido di Dante, whilst at the other locations pro-
or fishing on the structures varied with time of day
tection schemes covered up to more than 4 km. All the
(morning, midday and afternoon), state of the tide (low
structures were built with large blocks of quarried rock
and high), or day of the week (weekdays and week-
(mainly limestone), and set on shallow sediments, with
ends). For each study, sampling was replicated several
a maximum depth of about 3 m at the seaward sides of
times within each of winter, spring, summer and
breakwaters. People accessed structures either on foot,
autumn, in order to assess whether patterns observed
by swimming or by the use of little boats. While access
as a function of the above 3 factors were consistent
to groynes is allowed, the law bans access to break-
across season. Replicates consisted of 4 independent
countings of people on the structures (both break-
The structures supported assemblages dominated by
waters and groynes for a total length of about 900 m)
mussels (Mytilus galloprovincialis), oysters (Ostrea
on randomly chosen days representing the different
edulis and the non-indigenous Crassostrea gigas) and
conditions of season, tide, time of day and day of the
green algae, mainly Ulva intestinalis and the non-
week (see Table 2). Days of rough weather were
indigenous Codium fragile ssp. tomentosoides. Lim-
avoided, since preliminary surveys indicated that
pets (Patella caerulea) and crabs (including Pachy-
human activity would be scarce. A total of 112 obser-
grapsus marmoratus and Eriphia verrucosa) were also
vations were carried out during the 3 studies. The
common. The structures also provided a habitat for
a variety of fishes, some of which were targets for
Study 4 was carried out in May and August 2002 at 4
recreational anglers and spear fishermen (Table 1). A
locations selected at random (among 7 available) along
Table 1. Human recreational activities on and around coastal defence structures along the Emilia Romagna shores and principalorganisms harvested. Indication is given about whether the activity was legal or not, the prevailing habitat affected, the
prevailing time of the year of the activities, and their frequency
Harvesting of shellfish from the structures Musselsa Fishingf Angling Legalg Harvesting of clams from the sediments Striped venusi No-take activities Snorkelling
aMytilus galloprovincialisbOstrea edulis and Crassostrea gigascPatella caeruleadPachygrapsus marmoratus and Eriphia verrucosaePalaemon serratusf Main target species included Mugil spp., Lithognathus mormyrus, Boobs boops, Solea solea, Bothus spp., Trachinus draco,
Gobius paganellus and Dicentrarchus labrax
g These activities are legal. Access to breakwaters is, however, restricted by lawh Spear fishing on coastal defence structures is illegal during the bathing season (1 June to 31 September)iChamaelea gallinajSolen marginatus
Table 2. Designs of the 4 studies used to quantify people harvesting or fishing on coastal defence structures along the Emilia
Romagna shores and to assess spatial and temporal variation in such activities
Study 1: Lido di Dante, November 2001 to November 2002 Time of day
Morning (08:00 to 10:00 h), midday (12:00 to 14:00 h during autumn and winter or 13:00 to 15:00 h during spring and summer) and afternoon (14:00 to 16:00 h during autumn and winter or 16:00 to 18:00 h during spring and summer)
4 (counts of people on about 900 m of breakwaters and groynes over 2 h)
Study 2: Lido di Dante, November 2001 to November 2002 State of the tide
4 (counts of people on about 900 m of breakwaters and groynes over 2 h)a
Study 3: Lido di Dante, November 2001 to November 2002 Day of the week
4 (counts of people on about 900 m of breakwaters and groynes over 2 h)b
Study 4: 4 locations along the coast, May and August 2002 Location
Punta Marina, Lido Adriano, Lido di Savio and Cesenatico
5 (counts of people on about 300 m of breakwaters over 2 h)c
a In the North Adriatic Sea low tides occur in the afternoon over the autumn and winter and early in the morning over thespring and summer. Sampling was, therefore, done between 15:00 and 17:00 h during the autumn and winter and betweena06:00 and 08:00 h during the spring and summerb Sampling was done between 14:00 and 16:00 h during the autumn and winter and between 15:00 and 17:00 h during thebspring and summerc Sampling was done at low tide conditions between 06:00 and 08:00 h
Airoldi et al.: Impact of recreational harvesting
the coast: Punta Marina, Lido Adriano, Lido di Savio,
epibiota were thus quantified during the following
and Cesenatico. The study tested whether counts of
May (2004), when growth of species peaked and
people harvesting or fishing on the structures varied
human disturbance from the subsequent holiday sea-
among locations, and whether spatial patterns were
son was still relatively low. For each block, sampling
consistent between times of the year with different
was replicated in four 20 × 20 cm plots. Abundance (on
beach users. Preliminary surveys had suggested that
both primary and secondary substrata) of dominant
people visiting the shore in the spring were mainly
epibiota, and space not occupied by visible macro-
locals, while in the summer more tourists were present.
scopic organisms were quantified visually as percent
For this study, sampling was restricted to the breakwa-
covers (Benedetti-Cecchi et al. 1996). A frame divided
ters, as groynes were only present at Punta Marina.
into a grid of 25 sub-quadrats was used, and a score
Sampling was carried out early in the morning, and
from 0 to 4% was given to each taxon in each square.
was replicated 5 times in each of May and August.
Species present with <1% cover were given an arbi-
Each time, all locations were sampled by each of 4 ran-
domly allocated observers. Replicates were indepen-
The responses of epibenthic assemblages were
dent counts of people on about 300 m of structures.
examined by multivariate analyses on a total of 10
The period of observation for each replicate was 2 h.
response variables (see Results). Data were 4th-root
Data for the first 3 studies were analyzed using 2-way
transformed, to retain information regarding the rela-
ANOVAs, with season (4 levels) orthogonal to time of
tive abundance whilst at the same time reducing the
day (3 levels), tide (2 levels) or day of week (2 levels).
difference in scale among the variables (Clarke 1993).
Data from study 4 were analyzed using a 2-way
Principal coordinate analysis (PCO; Anderson 2003
ANOVA with location (5 levels) orthogonal to month
and references therein) was performed to produce a
(2 levels). Before running the analyses, data were
distance matrix of the centroids of each block in Bray-
logarithmically transformed, which always successfully
Curtis space, which was used as input to the sub-
stabilized variances (Cochran’s C-tests, p > 0.05).
sequent multivariate analyses. A plot of the first 2
Effects of mussel harvesting. An experiment was
principal coordinate axes was used to visualize pat-
carried out at Cesenatico to quantify the impact of
terns of overall dispersion of assemblages on blocks. In
mussel harvesting on dominant epibiota. It was de-
addition, a canonical analysis of principal coordinates
signed to test whether effects were consistent between
(CAP; Anderson & Willis 2003) was performed to visu-
the landward and seaward sides of breakwaters and
alize patterns of dispersion as a function of harvesting
between nearby breakwaters. The experiment was set
effects (using the a priori groupings of mussel harvest-
up on both the landward and seaward sides of each of
ing and control for the explanatory matrix) and side of
3 breakwaters, selected at random, about 100 to 600 m
breakwater (using the a priori groupings of landward
apart. For each side, 8 blocks about 1 m3, located from
and seaward sides). Effects were tested statistically
–20 to + 30 cm relative to mean-low-water and exten-
by using permutation 3-way multivariate analysis of
sively covered (> 80%) by mussels of harvestable size
variance (PERMANOVA, formerly NPMANOVA, An-
were permanently marked by securing stainless steel
derson 2001a), with harvesting (fixed, harvesting
bolts with marine epoxy into drilled holes. Blocks were
vs. control), side (fixed, landward vs. seaward) and
randomly allocated to either mussel harvesting (here-
breakwater (random, 3 levels) as orthogonal factors.
after ‘harvesting’) or unmanipulated (hereafter ‘con-
For the analyses, 4999 unrestricted random permuta-
trol’) treatments; there were 4 of each. Treatments
tions of the raw data were used to generate p-values
were applied in August 2003, as observations indi-
cated that this is the time of the year when harvesting
Responses of total macroalgae, mussels and space
is most intense. Mussels were removed from harvest-
not occupied by macroscopic organisms were further
ing blocks by using shovels, knives and chisels, in
examined by univariate 4-way ANOVAs. Factors were:
order to mimic as close as possible the observed
harvesting (fixed, harvesting vs. control), side (fixed,
changes caused by harvesters. Control blocks were left
landward vs. seaward) and breakwater (random, 3
untouched. The resulting harvested surfaces appeared
levels), orthogonal to each other, and block (random,
as patches of bare rock with sparse covers of byssal
4 levels) nested in the interaction of the previous 3
thread, and the remains of a few barnacles and macro-
factors. Pooling procedures were used when appropri-
ate to increase the power of the test for harvesting as in
In the study area, although some species settle late
Underwood (1997). The assumption of homogeneity of
in the summer or early autumn, recruitment and
variances was checked with Cochran’s C-tests, and
growth of species become generally visible in the
was always fulfilled (p > 0.05), except for unoccupied
spring (Bacchiocchi & Airoldi 2003, L. Airoldi et al.
space. Such heterogeneity could not be removed
unpubl. data). The effects of harvesting on the
through transformation of data; since the data set was
large and balanced, the analysis was done in any case
Spatial and temporal patterns
and results were interpreted conservatively, using p =0.01 as the level of significance (Underwood 1997).
A total of 145 people were observed harvesting
sessile organisms and 155 were observed fishing onthe coastal structures during the 304 h of observation
at our study sites. Most people were local males, butthe percentage of tourists and females increased over
Human activities and organisms affected
the summer. At Lido di Dante, where both groynesand breakwaters were present, over 70% of the peo-
Coastal structures were the target of recreational
ple preferentially exploited groynes, because they
activities, which involved the illegal exploitation of
were more easily accessible. The most illegal activi-
species (Table 1). Few people visited coastal defence
ties (e.g. collection of mussels for local restaurants,
structures, and particularly breakwaters, for recreational
fishing with nets or collection of large quantities of
no-take activities, probably because of poor accessibility
organisms) were, however, confined to the break-
and precarious footing on the steep and slippery surfaces
waters, and were often carried out at dawn or during
of the blocks. Mussels were by far the organisms most
the night. The reluctance of people to answer the
often harvested, followed by oysters, crabs, limpets and
questionnaire meant we could not identify clear pat-
shrimps (Table 1). Shellfish was mostly collected as food,
terns of shellfish harvesting as a function of position
although mussels and limpets were sometimes used as
on the structures. Our observations, however, sug-
bait for fishing. The following fishes, Mugil spp., Litho-
gest that mussel harvesting was most intense at the
gnathus mormyrus, Boops boops, Solea solea, Bothus
landward side of breakwaters at low shore levels,
spp., Trachinus draco, Gobius paganellus and Dicentrar-
while at the seaward side mussels were most often
chus labrax were among the most frequently caught
collected from shallow subtidal habitats (e.g. from
(Table 1). Interviews and direct observations suggested
that on average approximately 2.5 kg of mussels or 1 kg
The number of people harvesting shellfish on
of fish or crabs were collected per person each time.
groynes and breakwaters at Lido di Dante was consis-
Harvesting opened large patches within mussel beds,
tently greatest in summer, resulting in season having a
ranging from approximately 1 to several dm2.
significant effect in Studies 1 to 3 (Fig. 2, Table 3). Few
people were on the structures duringthe autumn and winter. During spring
some of the differences related totime of day, state of the tide and
differences were observed betweenlow and high tide in the summer
Fig. 2. Number of people harvesting shellfish on breakwaters and groynes dur-ing 4 seasons (W = winter, SP = spring, SU = summer, A = autumn) over 2001 and
2002 at Lido di Dante as a function of (a) time of day (Study 1), (b) tide (Study 2),
and (c) day of the week (Study 3); and (d) number of people harvesting shellfish
on breakwaters at 4 different locations (PM = Punta Marina, LA = Lido Adriano,
LS = Lido di Savio, and CS = Cesenatico) during May and August 2002 (Study 4).
Data are average counts over 2 h (+1 SE, n = 4 for Studies 1 to 3, n = 5 for
Study 4). For further details see Table 2
Airoldi et al.: Impact of recreational harvesting
Table 3. Results of ANOVAs and SNK tests on log-transformed numbers of people harvesting shellfish or fishing on coastal defencestructures during the 4 studies. Cochran’s C-tests for homogeneity was not always significant (p > 0.05). Significant p-values (p < 0.05)
are shown in bold type. Factors for each study are as in Table 2
Shellfish harvesting 1) Effect of < 0.01 < 0.01 < 0.01 Fishing 1) Effect of < 0.01
Morning: Summer = Spring > Autumn = Winter
Midday: Winter = Spring = Summer = Autumn
Afternoon: Spring = Autumn = Winter = Summer
< 0.01
waters among different locations along the Emilia
spring and summer than during the autumn and
Romagna coast (Fig. 2, Table 3, Study 4). This was
winter, resulting in season having a significant effect
unexpected; breakwaters were located at different dis-
in Studies 1 and 2 (Fig. 2, Table 3). No consistent
tances from the coast, thus exploitation might have
patterns in fishing activities were observed as a
been most intense at structures closest to the shoreline.
function of tide or day of the week (Fig. 3, Table 3,
Density of visitors was also similar between May and
Studies 2 and 3). Few differences were observed in the
August (Fig. 2, Table 3, Study 4), suggesting that
number of people fishing from the breakwaters at
mainly local people exploited breakwaters, whilst
different locations along the coast, both in May and
tourists made a minor impact, which is supported by
At Lido di Dante, numbers of recreational fishermen
were relatively constant across seasons at midday and
Effects of mussel harvesting on epibiota
in the afternoon, but varied in the morning (Fig. 3,Table 3, Study 1). Fishing was most frequent during
The variables included in the multivariate analyses
the morning in the summer and often in the spring,
were: Ulva intestinalis, filamentous algae, and Codium
while no fishing activity was recorded before midday
fragile spp. tomentosoides (subsequently lumped as
in the winter and autumn. Overall, there was a trend
‘macroalgae’ for univariate analyses as they showed
for a greater number of people to be fishing during the
similar responses to treatments), Mytilus galloprovin-cialis, oysters, unidentified sponges,
by a thin coating of unidentified micro-algae and/or other microorganisms,
a high proportion (often above 70%)of unoccupied space characterized
Fig. 3. Number of people fishing on breakwaters and groynes during 4 seasons
(W = winter, SP = spring, SU = summer, A = autumn) over 2001 and 2002 at Lido
di Dante as a function of (a) time of day (Study 1), (b) tide (Study 2) and (c) day of
the week (Study 3); and (d) number of people fishing on breakwaters at 4 differ-
ent locations (PM = Punta Marina, LA = Lido Adriano, LS = Lido di Savio, andCS = Cesenatico) during May and August 2002 (Study 4). Data are average
counts over 2 h (+1 SE, n = 4 for Studies 1 to 3, n = 5 for Study 4). For further
the experiment and before the newintensive harvesting season, assem-blages still differed significantly be-
tween harvested and control blocks. Differences were
particularly evident at the seaward sides of breakwa-
ters compared to the landward sides, which resulted ina significant interaction, harvesting × side (Table 4).
The significant differences found by the PER-
MANOVA test were not illustrated clearly in the prin-
cipal coordinate plot (Fig. 5a); this is likely because the
axis of real group differences that occurred in multi-
variate space was not in the same direction as the max-
imum variation. The plot of the 2 canonical axes corre-
sponding to the main effects of harvesting and side,
Table 4. Results of permutation multivariate analysis of vari-
ance (PERMANOVA) on Bray-Curtis dissimilarities for as-
semblages on breakwaters after 4th-root transformation.
Factors were: harvesting (fixed, harvesting vs. control), side
(fixed, landward vs. seaward) and breakwater (random, 3
levels). Significant effects (p < 0.05) are in bold
Fig. 4. Effects of experimental harvesting of mussels (in
August 2003) on covers of (a) total macroalgae (Ulva intesti-nalis, filamentous algae and Codium fragile spp. tomento-soides), (b) mussels (Mytilus galloprovincialis) and (c) un-
occupied space (bare rock, silt and a thin coating of
microorganisms) on the landward and seaward sides of each
of 3 manipulated and unmanipulated breakwaters at Cesen-
atico in May 2004. Data are average covers over 4 blocks per
breakwater and 4 quadrats per block (+1 SE, n = 16)
Airoldi et al.: Impact of recreational harvesting
Table 5. Results of ANOVAs on covers of total macroalgae (raw data), Mytilus galloprovincialis (raw data), and unoccupied space(log-transformed data). Factors are: breakwater (random, 3 levels), side (fixed, landward vs. seaward), harvesting (fixed, harvest-ing vs. control) and block (random, 4 levels, nested in the interaction of the other 3 factors). Cochran’s C-tests were not signifi-cant (p > 0.05) except for unoccupied space (p < 0.01). Significant effects (p < 0.05 for total macroalgae and M. galloprovincialis,
p < 0.01 for unoccupied space) are in bold. For further explanations see text
< 0.001 < 0.001 < 0.000
however, clearly showed a significant interaction
consistent between both the landward and seaward
sides of breakwaters and between breakwaters.
Differences between harvested and control blocks
The abundance of mussels in control blocks was
were further explained by univariate analyses on
lower than at the beginning of the experiment (Fig. 4),
macroalgae, Mytilus galloprovincialis, and unoccupied
because violent storms during the autumn and winter
space. Oysters, sponges and barnacles were too sparse
struck the breakwaters and extensively damaged the
for further meaningful analyses. Macroalgae flour-
mussel beds (authors’ unpubl. data). Their effects on
ished on the breakwaters, particularly on the seaward
mussels were particularly severe at the landward side
exposed sides, with peak covers above 60% (Fig. 4).
of the breakwaters, and probably masked some of the
Macroalgal cover expanded when mussels were har-
effects of recreational harvesting (see ‘Discussion’). By
vested, with average values of 39% in harvested
May, cover of mussels at harvested and control blocks
blocks and 25% in control blocks (Fig. 4). This increase
on the landward sides of the structures had reached
was detected as significant despite the variability in
comparable values (Fig. 4). On the seaward sides
macroalgal cover between blocks (Table 5), and was
mussels were on average more abundant in control
Fig. 5. Two-dimensional plots of the (a) principal coordinate axes (unconstrained metric multi-dimensional scaling) and(b) canonical axes of principal coordinates showing ordination of centroids of assemblages on breakwaters in each combinationof harvesting vs. control treatment and landward vs. seaward sides (n = 12, 4 blocks for each of 3 breakwaters). Analyses were
based on Bray-Curtis dissimilarities after 4th-root transformation
(54% cover) than in harvested (41% cover) blocks, but
possibly compensating the major efforts required to
patterns were variable over space and this difference
The estimates of sessile species, fish and crabs
At the landward sides of breakwaters there was a
collected per person, along with the counts of total
large amount of unoccupied space, much of it coated
harvesters, clearly demonstrate the dramatic levels of
with silt and/or microorganisms. This unoccupied
disturbance to assemblages on coastal structures in
space was produced both by the experimental harvest-
this region. For example, at Lido di Dante, during the
ing of mussels and by the subsequent effects of storms.
summer, an average of 3 people were observed har-
By May, unoccupied space was less abundant in har-
vesting shellfish on the structures every 2 h from
vested than control blocks (Fig. 4), probably as a con-
08:00 to 18:00 h, independently of time of day, day of
sequence of the growth of macroalgae in harvested
the week or tidal conditions (Fig. 2), which gives an
treatments. Although at the seaward sides of break-
estimate of 15 people d–1. Considering that about 80%
waters the amount of unoccupied space was small, a
of those people collected mussels, and that on aver-
similar trend was observed, which resulted in a sig-
age each person harvested about 2.5 kg of mussels,
nificant main effect of the harvesting factor (Table 5).
we estimate that ~2.7 tons of mussels are collectedover each summer on the coastal structures at Lido diDante alone. Measures of mussel biomass per unit
DISCUSSION
surface in June 2003 (F. Bacchiocchi unpubl. data)indicate an average wet weight of 188 g dm2 (n = 70,
The present study shows that disturbance from
SD = 104). Thus, 2.7 tons of harvested mussels would
exploitation has large effects on assemblages of coastal
roughly correspond to a disrupted surface of ~144 m2.
defence structures, and should be taken into account
This estimate is limited to a band of ~20 cm in height
when explaining their structure and variability. As-
across low shore and shallow subtidal habitats, where
semblages on these and other types of artificial struc-
mussels are an edible size and easy to collect; this
tures are, in fact, shaped and dependent on human
does not include people collecting mussels overnight
interventions; ignoring human perturbations in such
systems could lead to misleading conclusions about
Harvesting of mussels over the summer led to deple-
tion of mussel beds, opening of unoccupied space and
Recreational exploitation was a major and recurrent
patchiness in the assemblages, ultimately favouring
source of disturbance for the assemblages on defence
the development of green and filamentous algae,
structures along the highly populated coasts of Emilia-
including the invasive Codium fragile ssp. tomento-
Romagna. Hundreds of people visited the structures
soides. The effects of mussel harvesting were still
for recreational fishing and to harvest a variety of
notable after almost 1 yr from the beginning of the
invertebrates to be used as food or, to a lesser extent,
experiment. Differences were marked on the seaward
as bait. A legal ban did not act as a deterrent, rather it
sides of breakwaters; on the landward sides, effects
confined the most illegal activities to dawn or night.
were partially masked by the action of storms during
Different from rocky shores in other parts of the world
the autumn and winter, which damaged mussel beds
(e.g. Kingsford et al. 1991, Addessi 1994, Keough &
on the structures. Damage was particularly severe on
Quinn 1998), few people visited the structures, and
the landward side of breakwaters compared to the sea-
particularly breakwaters, for recreational no-take
ward sides, probably because mussel beds at these
sheltered habitats were multi-layered, thus more sub-
Human exploitation was most intense during the
ject to the risk of dislodgement by waves (Harger &
spring and summer, and activities that required
Landenberger 1971, Paine & Levin 1981). It is also pos-
immersion in the water, such as mussel harvesting,
sible that mussel beds were particularly susceptible to
were restricted to the warmest months. Density of visi-
storms after being weakened by intensive harvesting;
tors varied at small temporal scales, but relationships
the compactness of mussel beds can, in fact, be com-
with time of day, state of the tide or day of the week
promised by the removal of even a few individuals
were not obvious. Exploitation was evenly distributed
(Denny 1987), and a number of small disturbed points
among different locations along the coast, despite
may coalesce to produce larger patches (Paine & Levin
marked differences in the accessibility of the struc-
1981). A ‘human-exclusion’ experiment (e.g. Castilla
tures. Distance from the shore was probably not a
1999) would be necessary to test whether human har-
limiting factor for local people, who were the main
vesting can enhance the susceptibility of mussel beds
exploiters. Furthermore, breakwaters more distant
to wave dislodgement. We attempted such an experi-
from the shore are an ideal place to carry out unlawful
ment in 2002. As legislative protection was lax, the
activities since they are more difficult to observe, thus
experiment had to rely on the voluntary abstention of
Airoldi et al.: Impact of recreational harvesting
people from harvesting at certain sites. Despite the
Acknowledgements. This work was supported by the EU
use of explanatory signs at both the breakwaters
project DELOS (EVK3-CT-2000-00041) and by the projectCOFIN (ex 40%). L.A. was supported by an Assegno di
and beach establishments, people’s collaboration was
Ricerca of the University of Bologna. We thank J. M. Ander-
minimal, and the experiment was abandoned.
son, F. Bertasi, M. Carrera, F. Colosio, F. Costantini and G. M.
The exploitation of assemblages on man-made
Branca for help with the fieldwork, and 4 anonymous review-
structures as sources of harvestable biomass of macro-
ers for their careful revision of the manuscript. We are grate-
algae, shellfish and fishes is also a common phenome-
ful to T. J. Willis for help with the statistical analyses and forinsightful comments on an earlier draft.
non in other coastal areas (Collins et al. 1994). Ourresults emphasize the need for effective managementof such human disturbances to urban coastal struc-
tures. These habitats attract ‘artificial’ assemblages
Abelson A, Shlesinger Y (2002) Comparison of the development
that are often extraneous to the native characteristics
of coral and fish communities on rock-aggregated artificial
of the areas and that can disturb regional species
reefs in Eilat, Red Sea. ICES J Mar Sci 59:S122–S126
diversity and introduce non-native or nuisance spe-
Addessi L (1994) Human disturbance and long-term changes
cies (Glasby & Connell 1999, Lambert & Lambert
on a rocky intertidal community. Ecology 4:786–797
2003, Airoldi et al. 2005). Thus, any factor influencing
Airoldi L, Abbiati M, Beck MW, Hawkins SJ and 6 others
(2006) An ecological perspective on the deployment and
the type and distribution of organisms on man-made
design of low-crested and other hard coastal defence
structures requires careful consideration of its ulti-
mate effects on native environments. For example, the
Anderson MJ (2001a) A new method for non-parametric
proliferation of hard coastal structures along the
multivariate analysis of variance. Austral Ecol 26:32–46
Anderson MJ (2001b) Permutation tests for univariate or
sandy coasts of Emilia Romagna has promoted the
multivariate analysis of variance and regression. Can J
expansion of a number of extraneous species, includ-
ing the pest alga Codium fragile ssp. tomentosoides
Anderson MJ (2003) PCO: a FORTRAN computer program for
(Bulleri et al. 2005, Bulleri & Airoldi 2006). Further,
principal coordinate analysis. Department of Statistics,
flourishing of algae on the structures is a problem
Anderson MJ, Willis TJ (2003) Canonical analysis of principal
for local tourism. Algae are torn off and wash onto
coordinates: a useful method of constrained ordination for
the beaches, thereby reducing the beaches perceived
amenity; they also need to be removed periodically,
Ardizzone GD, Belluscio A, Gravina MF, Somaschini A (1996)
this adds costs to local municipalities. Although the
Colonization and disappearance of Mytilus galloprovin-cialis Lam. on an artificial habitat in the Mediterranean
growth of Codium and other algae is an unavoidable
consequence of the presence of the structures, distur-
Artegiani A, Bregant D, Paschini E, Pinardi N, Raicich F,
bance to mussel beds indeed facilitates their growth.
Russo A (1997) The Adriatic Sea general circulation.
Limiting human exploitation would help control the
Part II. Baroclinic circulation structure. J Phys Oceanogr
abundance and spread of these nuisance species;
Bacchiocchi F, Airoldi L (2003) Distribution and dynamics of
thus, there are substantial environmental and socio-
epibiota on hard structures for coastal protection. Estuar
economic reasons why an effective regulation of
human visits to the structures would be desirable.
Benedetti-Cecchi L, Airoldi L, Abbiati M, Cinelli F (1996)
Clearly, the management of recreational activities
Estimating the abundance of benthic invertebrates: a com-parison of procedures and variability between observers.
on defence structures along the coasts of Emilia Ro-
magna has not received appropriate attention, and
Boalch GT, Holme NA, Jephson NA, Sidwell JMC (1974) A
the almost complete absence of visible enforcement
resurvey of Colman’s intertidal traverses at Wembury,
contributes to the high frequency of unlawful visita-
South Devon. J Mar Biol Assoc UK 54:551–553
tions. Advice that recreational activities should be
Bulleri F, Airoldi L (2006) Artificial structures as habitats
for non-indigenous species: the case of the green alga,
regulated more effectively has also been proposed for
Codium fragile ssp. tomentosoides, in the north Adriatic
natural rocky shores (e.g. Fanelli et al. 1994, Murray
Sea (North-eastern Mediterranean). J Appl Ecol (in press)
et al. 1999, Keough & Quinn 2000). Although the
Bulleri F, Chapman MG (2004) Intertidal assemblages on arti-
nature, scale and consequences of recreational ex-
ficial and natural habitats in marinas on the north-westcoast of Italy. Mar Biol 185:381–391
ploitation substantially differ between man-made and
Bulleri F, Abbiati M, Airoldi L (2005) The colonisation of arti-
natural hard substrata, human harvesting is a major
ficial human-made structures by the invasive alga Codium
disturbance for coastal assemblages in both habitats,
fragile ssp. tomentosoides in the north Adriatic Sea (NE
affecting their distribution and structure. Knowledge
of the type, magnitude and variability of such distur-
Castilla JC (1999) Coastal marine communities: trends and
perspectives from human-exclusion experiments. Trends
bances and recognition of their ecological effects can
help to develop policies for managing human ex-
Cencini C (1998) Physical processes and human activities in
the evolution of the Po delta, Italy. J Coast Res 14:774–793
Chapman MG (2003) Paucity of mobile species on con-
Guidetti P, Bussotti S, Boero F (2005) Evaluating the effects of
structed seawalls: effects of urbanization on biodiversity.
protection on fish predators and sea urchins in shallow
artificial rocky habitats: a case study in the northern
Chapman MG, Bulleri F (2003) Intertidal seawalls — new
Adriatic Sea. Mar Environ Res 59:333–348
feautures of landscape in intertidal environements. Land-
Harger JRE, Landenberger DE (1971) The effect of storms
as a density dependent mortality factor on populations of
Choi CG, Takeuchi Y, Terawaki T, Serisawa Y, Ohno M, Sohn
CH (2002) Ecology of seaweed beds on two types of artifi-
Keough MJ, Quinn GP (1998) Effects of periodic disturbances
from trampling on rocky intertidal algal beds. Ecol Appl 8:
Clarke KR (1993) Non-parametric multivariate analyses of
changes in community structure. Aust J Ecol 18:117–143
Keough MJ, Quinn GP (2000) Legislative vs. practical protec-
Collins K, Jensen A, Lockwood A, Lockwood S (1994) Coastal
tion of an intertidal shoreline in southeastern Australia.
structures, waste materials and fishery enhancement. Bull
Kingsford MJ, Underwood AJ, Kennelly SJ (1991) Humans as
Connell SD (2001) Urban structures as marine habitats: an
predators on rocky reefs in New South Wales, Australia.
experimental comparison of the composition and abun-
dance of subtidal epibiota among pilings, pontoons and
Lambert CC, Lambert G (2003) Persistence and differential
rocky reefs. Mar Environ Res 52:115–125
distribution of nonindigenous ascidians in harbors of the
Connell SD, Glasby TM (1999) Do urban structures influence
Southern California Bight. Mar Ecol Prog Ser 259:145–161
local abundance and diversity of subtidal epibiota? A case
Lindberg DR, Estes JA, Warheit KI (1998) Human influences
study from Sydney Harbour, Australia. Mar Environ Res
on trophic cascades along rocky shores. Ecol Appl 8:
Davis JLD, Levin LA, Walther SM (2002) Artificial armored
Majiza VN, Lasiak TA (2002) The influence of site, season and
shorelines: sites for open-coast species in a southern
day of the week on exploitation of rocky intertidal biota in
central Transkei, South Africa. S Afr J Mar Sci 24:57–64
De Boer WF, Blijdenstein AF, Longamane F (2002) Prey
Meinesz A, Lefevre JR, Astier JM (1991) Impact of coastal
choice and habitat use of people exploiting intertidal
development on the infralittoral zone along the south-
eastern Mediterranean shore of continental France. Mar
Denny MW (1987) Lift as a mechanism of patch initiation in
mussel beds. J Exp Mar Biol Ecol 113:231–245
Milazzo M, Badalamenti F, Riggio S, Chemello R (2004) Pat-
Dethier MN, McDonald K, Strathmann RR (2003) Coloniza-
terns of algal recovery and small-scale effects of canopy re-
tion and connectivity of habitat patches for coastal marine
moval as a result of human trampling on a Mediterranean
species distant from source populations. Conserv Biol 17:
rocky shallow community. Biol Conserv 117:191–202
Moreno CA (2001) Community patterns generated by human
Durán LR, Castilla JR (1989) Variation and persistence of the
harvesting on Chilean shores. Aquat Conserv 11:19–30
middle rocky intertidal community of central Chile, with
Moschella PS, Abbiati M, Åberg P, Airoldi L and 12 others
and without human harvesting. Mar Biol 103:555–562
(2005) Low crested coastal defence structures as artificial
Dye AH (1992) Experimental studies of succession and stabil-
habitats for marine life: using ecological criteria in design.
ity in rocky intertidal communities subject to artisanal
shellfish gathering. Neth J Sea Res 30:209–217
Murray SN, Denis TG, Kido JS, Smith JR (1999) Human visi-
Fanelli G, Piraino S, Belmonte G, Geraci S, Boero F (1994)
tation and the frequency and potential effects of collecting
Human predation along Apulian rocky coasts (SE Italy):
on rocky intertidal populations in southern California
desertification caused by Lithophaga lithophaga (Mol-
marine reserves. CalCOFI Rep 40:100–106
lusca) fisheries. Mar Ecol Prog Ser 110:1–8
Page HM, Dugan JE, Dugan DS, Richards JB, Hubbard DM
Forni S, Malagoli M (2004) Rapporto annuale sul movimento
(1999) Effects of an offshore oil platform on the distri-
turistico e la composizione della struttura ricettiva (al-
bution and abundance of commercially important crab
berghiera e complementare) dell’Emilia Romagna: anno
2003. Regione Emilia Romagna, Servizio Turismo e Qual-
Paine RT, Levin SA (1981) Intertidal landscapes: disturbance
ità Aree Turistiche della Regione, Bologna
and the dynamics of patterns. Ecol Monogr 51:145–178
Fraschetti S, Bianchi CN, Terlizzi A, Fanelli G, Morri C, Boero
Rebele F (1994) Urban ecology and special features of urban
F (2001) Spatial variability and human disturbance in
ecosystems. Global Ecol Biogeogr 4:173–187
shallow subtidal hard substrate assemblages: a regional
Southward AJ, Orton JH (1954) The effects of wave-action on
the distribution and numbers of the commoner plants and
Glasby TM (1999) Differences between subtidal epibiota on
animals living on the Plymouth breakwater. J Mar Biol
pier pilings and rocky reefs at marinas in Sydney. Estuar
Underwood AJ (1997) Experiments in ecology: their logical
Glasby TM, Connell SD (1999) Urban structures as marine
design and interpretation using analysis of variance. Editorial responsibility: Otto Kinne (Editor-in-Chief),Submitted: November 19, 2004; Accepted: March 31, 2005Proofs received from author(s): July 29, 2005
T h i s p a g e a n d i t s c o n t e n t s M I R E N A C O I L F O R H E A V Y P E R I O D S t h e B r i t i s h F i b r o i d T r u s tDr Nicki On, PhD, MRPharmS. Pharmacist Dr Rajesh Varma, MA, PhD, MRCOG. Consultant Obstetrician & Gynaecologist. Website address: www.britishfibroidtrust.org.uk 1. An IUS (intrauterine system), commonly know as a coil, is a small T-shaped plastic d
Diagnostiek 1. Audiogram (gehoortest): Doel is vaststellen van gehoorsverlies in decibel. U krijgt een koptelefoon op waaruit afwisselend uit de linkerkant en de rechterkant piepjes van verschillende toonhoogte te horen zijn. Wanneer u een piepje hoort geeft u dat aan. De assistente kan zo in kaart brengen welke toonhoogte u op welke sterkte hoort. Het resultaat wordt besproken met de
 Airoldi et al.: Impact of recreational harvesting
description of the assemblages can be found inBacchiocchi & Airoldi (2003) and Bulleri et al. (2005).
Patterns of human activities. The types of recre-
Airoldi et al.: Impact of recreational harvesting
description of the assemblages can be found inBacchiocchi & Airoldi (2003) and Bulleri et al. (2005).
Patterns of human activities. The types of recre-