Le métronidazole (Flagyl) reste la référence dans le traitement des infections anaérobies et des parasitoses comme la giardiase ou l’amibiase. Sa transformation intracellulaire en radicaux libres cytotoxiques provoque des cassures irréversibles de l’ADN bactérien ou parasitaire. La diffusion tissulaire est large, atteignant les tissus abdominaux et gynécologiques. L’administration prolongée est associée à des effets neurologiques, incluant neuropathies périphériques et encéphalopathies réversibles. L’association avec l’alcool déclenche une réaction de type antabuse. Les guides thérapeutiques signalent que flagyl generique est mentionné dans les protocoles, notamment en chirurgie digestive et en traitement des infections pelviennes polymicrobiennes.
Jm520507 403.408
Journal of Medical Microbiology (2003), 52, 403–408
Expression stability of six housekeeping genes: aproposal for resistance gene quantification studiesof Pseudomonas aeruginosa by real-timequantitative RT-PCR
Hakan Savli,1 Aynur Karadenizli,2 Fetiye Kolayli,2 Sibel Gundes,3Ugur Ozbek4 and Haluk Vahaboglu3
1–3Tibbi Biyoloji AD1, Mikrobiyoloji ve Klinik Mikrobiyoloji AD2 and Enfeksiyon Hastaliklari ve Klinik
Mikrobiyoloji AD3, Kocaeli Universitesi, Tip Fakultesi, Sopali Ciftligi, 41900 Kocaeli, Turkey
4Genetik AD, DETAE, Istanbul Universitesi, Istanbul, Turkey
Constantly expressed genes are used as internal controls in relative quantification studies. Suitableinternal controls for such studies have not yet been defined for Pseudomonas aeruginosa. In thisstudy, the genes ampC, fabD, proC, pbp-2, rpoD and rpoS of P. aeruginosa were compared in termsof expression stability by real-time quantitative RT-PCR. A total of 23 strains with diverse resistancephenotypes were studied. Stability of expression among the housekeeping genes was assessed onthe basis of correlation coefficients, with the best-correlated pair accepted as being the most stableone. Eventually, proC and rpoD formed the most stable pair (r ¼ 0·958; P , 0·001). Next, in fourciprofloxacin-selected nfxC-like mutants, levels of oprD, oprM and oprN mRNA were compared withthose of their wild-type counterparts. The comparison was made after correcting the raw values bythe geometric mean of the internal control genes proC and rpoD. The level of oprN mRNA wassignificantly up-regulated, while the oprD gene was down-regulated (although this difference wasstatistically insignificant), in the mutants. This expression pattern was consistent with that of the
expected expression profile of nfxC-type mutants; this experiment therefore lends further support to
the use of proC and rpoD genes simultaneously as internal controls for such studies.
Levels of expression of these genes have traditionally beenstudied by Western blotting with the aid of mAbs. Unfortu-
Pseudomonas aeruginosa is an important nosocomial patho-
nately, this method is unable to help in studying different
gen, particularly in intensive care units 2000).
proteins simultaneously and so is weak in relative compari-
Members of this species are inherently resistant to a range of
sons. Moreover, mAbs are not commercially available. The
antibiotics. In addition, they are capable of conferring
quantification of mRNAs by real-time RT-PCR has been
resistance to others by shifting the regulation levels of various
used with great success in other fields , 2000;
, 1999). We suggest that this highly sensitivemethod may also be useful in relative comparison of
Porins (particularly porin D) and efflux pumps of the
resistance-nodulation-division (RND) family are the cur-
The reliability of a relative comparison depends largely on the
rently recognized genetic systems that are associated sig-
normalization of unwanted variations between samples.
nificantly with antibiotic resistance , 2001).
Constantly expressed genes, often selected from among
The exact links between the expression patterns of these and
housekeeping genes, are used as internal controls for normal-
other resistance phenotypes have not been fully elucidated,
ization of the results. The proportion of mRNA of constantly
and inconsistent conclusions appear in the literature
expressed genes in the total cellular RNA is assumed to be
equal between different samples. Normalization of raw
values by means of internal controls therefore serves to
topic still attract considerable interest.
eliminate sample-to-sample variation of the RNA isolationand reverse transcription steps and, even more importantly,serves to eliminate variations in total transcriptional activity
For this purpose, six housekeeping genes were compared in
39 8C (with an increase of 0·3 8C every cycle), 2 min at 72 8C and 1 min
this study. These were pyrroline-5-carboxylate reductase
at 94 8C and 30 cycles of 2 min at 44 8C, 3 min at 72 8C and 1 min at
(proC), malonyl CoA : acyl carrier protein (ACP) transacyl-
94 8C. A final extension for 1 h at 72 8C completed the procedure.
ase (fabD), sigma factors RpoD (rpoD) and RpoS (rpoS),
PCR products were separated on a 2 % agarose gel and visualized by
penicillin-binding protein 2 (pbp-2) and chromosomal beta-
ethidium bromide staining. For better resolution, they were also run on
lactamase (ampC). The expression stability of these house-
a 6 % acrylamide/bis-acrylamide gel (data not shown). The banding
keeping genes was first investigated as proposed by
patterns of both gels were analysed by the freely distributed gel analysis
et al. (2002) and, later, the levels of oprM,
software LabImage (version 2.62). Molecular sizes of the bands werecalculated by this software relative to the marker DNA.
oprN and oprD mRNA were compared in a set of P. aeruginosa strains.
RNA isolation and reverse transcription. Total RNA was isolatedfrom 5 ml fresh overnight (approx. 18 h) broth culture (MH broth) byusing the NucleoSpin RNA II kit (Macherey-Nagel), as described by the
manufacturer. Genomic DNA was eliminated by RNase-free DNase Itreatment during the isolation procedure. Finally, RNAs were eluted
Strains, resistance tests and beta-lactamase assays. We selected
from the silica membranes in a volume of 40 ìl diethyl pyrocarbonate-
17 P. aeruginosa strains from a set obtained from university hospitals in
treated double-distilled water. The A260 of the resulting RNA solution
four different geographical regions of Turkey. The bacteria had been
was between 1 and 10. Reverse transcription was performed at 42 8C for
identified in those universities by various identification systems. The
90 min by using random hexamer primers so as to obtain cDNA copies
strains were, nevertheless, reidentified in our institute by classical
of mRNAs (2 ìl) with 100 IU MMuLV reverse transcriptase (MBI
methods and, if required, by the non-fermenter ID panel of the Becton
Fermentas) in 20 ìl total volume. Concentrations of cDNAs were
Dickinson system (Diagnostic Instrument Systems).
adjusted on a LightCycler (Roche Diagnostics). For every sample, 1 ìlcDNA and 9 ìl SYBR Green I (the same concentration as indicated by
Of the 17 strains selected, 10 were fully susceptible to antibiotics of the
the manufacturer for the PCR assay) were mixed in capillary tubes. After
main classes, while the others were of different resistance phenotypes.
incubation at 95 8C for 5 min, fluorescence emissions were read at 55 8C
We challenged the susceptible strains in Mueller–Hinton (MH) broth
with the real-time fluorometry facility of the LightCycler. This enabled
with ciprofloxacin (0·1 ìg mlÀ1) in order to obtain resistant variants.
us to compare the total cDNA concentrations of the samples with the
Consequently, six isogenic mutants with various resistance patterns
control transcript, which was approximately 1 mg mlÀ1 at the highest
were selected. These mutants, as well as their counterparts, were
dilution. Concentrations of cDNAs of the samples were adjusted to a
level close to the second dilution (10À1) of the control cDNA. Thisadjustment was critical for performing successful calculations. The aim
Resistance patterns were determined by the disk diffusion method on
was to keep the cDNA concentrations of the samples between the
MH agar plates. MICs were determined by either E-test strips on MH
concentrations of the controls in order to avoid large variations during
agar plates or by the agar dilution method as described by the NCCLS.
calculations by the LightCycler software.
Antibiotic disks and MH agar were obtained from Oxoid while E-teststrips were sourced from AB Biodisc. Powder forms of the antibiotics
Real-time PCR. The sequences of the genes studied were obtained from
were obtained as gifts from the respective companies.
GenBank and the primers were designed with the aid of the OLIGOsoftware (version 5.0; Molecular Biology Insights). The sequences of the
Beta-lactamases were analysed as described elsewhere
1998). Extracts obtained by freezing and thawing of dense bacterialsuspensions were applied to polyacrylamide gels with ampholytes
PCR was performed in the LightCycler in capillary glass tubes with the
ranging from pH 3 to 10. Nitrocefin overlay and migration relative to
LightCycler FastStart DNA Master SYBR Green I kit (Roche). Work was
TEM-1 and SHV-1 standards enabled us to evaluate the pI values of
always carried out on desktop coolers (pre-cooled to 4 8C). Master
beta-lactamases. Precise identification of the beta-lactamases depended
mixtures were prepared exactly as recommended by the manufacturer,
on sequence analysis of PCR products as described previously
except for the concentration of Mg2þ. The final concentrations of Mg2þ
and primers were respectively 2·5 mM and 50 pmol per reaction.
The control cDNA was from P. aeruginosa ATCC 27853 and the primers
Random amplified polymorphic DNA (RAPD) typing. Clonal
of the control reactions were specific for pbp-2. An arbitrary concentra-
variability was further ensured by RAPD typing of eight selected strains,
tion value of 1·5 3 104 copies of the pbp-2 gene was assigned to the
two from each region. DNA was isolated from fresh overnight agar
control transcript. Tenfold dilutions of this down to 15 copies of the
cultures. A loopful of bacteria was homogenized in 50 ìl TE buffer (10
pbp-2 gene were always included in the reactions.
mM Tris/HCl, 1 mM EDTA, pH 8) and lysis was accomplished with500 ìl guanidium thiocyanate (6 M) plus 0·5 % sodium lauroylsarco-
PCR was accomplished after a 5 min activation and denaturation step at
sine for 10 min at room temperature. DNA was extracted first by
95 8C, followed by 45 cycles of 15 s at 95 8C, 10 s at 60 8C and 15 s at
phenol/chloroform and then by chloroform/isoamyl alcohol (24 : 1,
v/v) and then precipitated with 0·1 vols sodium acetate (3 M, pH 5·4)
Primer dimers and other artefacts were evaluated by melting curve
plus an equal volume of 2-propanol at room temperature. Precipitates
analysis and eventually only dimer- and artefact-free reactions were
were collected by centrifugation (10 min at 12 000 g) and then the pellets
considered valid. Results were read with the ‘second derivative maxi-
were washed twice with 70 % ethanol, air-dried for 2 min and
mum’ algorithm of the software provided. The LightCycler software
resuspended in 30 ìl double-distilled water.
generated a standard curve by plotting ‘crossing cycle number’ versus
RAPD PCR was performed as described elsewhere
logarithms of the given concentrations for each control. Eventually, a
regression line was drawn between these points. The software calculated
GGATTCA-39) and ERIC-2 (59-AAGTAAGTGACTGGGGTGAGCG-
the concentrations of the studied genes with the aid of this standard
39). Master mixtures were prepared as described in the above references.
However, the amplification program was modified as follows: one cycleof denaturation for 5 min at 95 8C followed by 25 cycles of 3 min at
Statistical analysis. The stability of mRNA expression was assessed by
Housekeeping genes of Pseudomonas aeruginosa
Table 1. Primers used in quantification studies
calculation of Spearman’s correlation coefficients of the raw concentra-tion data with the aid of the statistical package SPSS (version 9.0). The
best-correlated pair was considered to be the most stable one.
Stability was further evaluated by a freely distributed MS Excelapplication (geNorm). Detailed information on this application can
be obtained al. (2002). This approach assumesthat minimally regulated, stably expressed genes stay in a constant ratio
to each other. In other words, in a given set of genes, it must be the pair ofmost stable genes that will be able to keep the ratio to each otherconstant in different samples. Importantly, co-regulated genes areexceptions to this assumption and they are not included.
The applet geNorm helps to calculate the gene expression stabilitymeasure (M), which is the mean pair-wise variation for a gene from all
other tested control genes al., 2002). A higher value ofM means greater variation in expression. The stepwise exclusion ofgenes with the highest M values allows the ranking of the tested genesaccording to their expression stability. The proposed threshold foreliminating a gene as unstable was an expression stability measure of> 0·5.
Raw quantities were corrected by dividing a value by the geometricmean of proC and rpoD genes of the same sample. Relative comparisonswere done between corrected values with the ANOVA test for
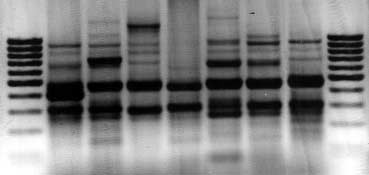
Fig. 1. (a) RAPD fingerprints (negative exposure). Lanes: M, mole-
cular size marker (100 bp DNA ladder); 1–7, selected strains (strains2 and 5–7 are the parents of nfxC mutants). (b) Fingerprints
A total of 23 strains were included in the study. The group
according to the lengths (bp) of fragments calculated by gel analysis
was composed of 17 wild-type strains and six ciprofloxacin-
software. Lanes are the same as in (a).
selected, isogenic mutants. Clonal heterogeneity was inves-tigated by DNA typing of eight of these wild-type strains. Onesample did not provide a readable pattern, probably becauseof inhibitors carried over from the DNA isolation step.
nfxC mutants because of the concurrent down-regulation of
However, the other seven were amplified successfully and the
fingerprints were sufficiently polymorphic to confirm the
phenotypes of the mutants were in agreement with this.
Interestingly, the MICs of ceftazidime were variable; one was
Of the 17 wild-type strains, seven were already resistant to
increased and one decreased while the other two were
various antibiotics. It is noteworthy that three of these did
unchanged. One study reported an unexplained decrease in
not produce a beta-lactamase other than the chromosomal
the ceftazidime MICs of nfxC mutants 2000).
one strains were probably resistant because
However, another study showed that the functional subunit
of activated porin and/or efflux systems. Of the six cipro-
of the mexEF–oprN operon is not related to beta-lactam
floxacin-selected mutants, two were resistant only to the
hypersusceptibility 2000). These observations
fluoroquinolones, while the other four expressed nfxC-like
suggest the existence of other co-operating systems respon-
multiple-resistance phenotypes In addition to
sible for beta-lactam susceptibility or resistance in nfxC
fluoroquinolones, the MIC of imipenem increases among
mutants. This issue remains unresolved.
Table 2. Antibiotic-resistance phenotypes and extrinsic beta-lactamases in strains examined
Seventeen strains were examined: strains R1–7, resistant wild-type; S1–10, susceptible wild-type. Strains were classed asresistant (R) or susceptible (S) according to NCCLS criteria.
*Other than chromosomal AmpC beta-lactamase.
Table 3. MICs of ciprofloxacin-selected (nfxC-like) mutants compared with wild-types
MICs are given in ìg mlÀ1 for the parent strain/mutant.
The housekeeping metabolism of prokaryotes has been
These data led us to assume that these genes might also be
shown to be highly variable et al., 2001), so
expressed in sufficient quantities in P. aeruginosa, a further
genes expressed stably under one condition might not be so
advantage in optimization of the PCR test.
under others. Stability in terms of mRNA expression inprokaryotic cells, therefore, should be tested under equiva-
The raw quantities of mRNA of the six genes studied
lent conditions with the investigated setting. In other words,
internal controls intended for use in resistance gene quanti-
coefficients indicated proC and rpoD as the most significant
fication studies should be tested in strains with changing
pair (r ¼ 0·958; P , 0·001). Similarly, the stepwise exclusion
resistance phenotypes. The diversity in the resistance phe-
of the genes with the highest M values by geNorm left proC
notypes of this study group fulfils this requirement.
(M ¼ 0·36) and rpoD (M ¼ 0·36) as the most stable genes.
Next, the levels of oprD, oprM and oprN mRNA of the fournfxC-like isogenic mutants were compared with those of
Six genes were compared in this study group. In order to
their wild-type counterparts. In this experiment, the geo-
avoid co-regulated genes, we carefully selected genes that are
metric mean of the levels of proC and rpoD in a sample was its
distantly related in metabolic function and chromosomal
normalization factor. Comparison of the normalized quan-
order. The selected genes participate in critical functions.
titative values of these genes is shown mean
pbp-2 has a central role in peptidoglycan metabolism, while it
level of oprN was 1·3 times higher in the mutants, while the
has some relation to the rod-shape-determining protein.
mean level of oprD was 1·28 times lower. Values were
comparable for all the mutants. Interestingly, the statistical
The sigma factor-encoding rpoD is a critical housekeeping
comparison was significant only for oprN concentrations.
gene , 1995). proC is involved in amino acidbiosynthesis, while fabD is involved in a different class of
Data on expression of these resistance genes in the literature
metabolic function , 1999). However, their
have to date been obtained by immunoblotting. Evaluations
metabolic importance was not the only reason for selecting
were dependent on crude differences and lacked precise
these genes. Equally important is that they all were shown in
numerical values. Therefore, comparison of the results of this
an Escherichia coli DNA array study to be expressed at levels
study with the literature was not possible. Moreover,
immunoblotting indicates differences in the level of mature
Housekeeping genes of Pseudomonas aeruginosa
Table 4. Raw concentrations of mRNA of six genes from 23 strains obtained by RT-PCR
Concentrations are given as copies ìlÀ1 (310À3). S1–S10 are susceptible strains; S5 and S6 are theparents of M1 and M2 (ciprofloxacin-selected non-nfxC-type mutants) and S7–S10 are theparents of NFX1–NFX4 (ciprofloxacin-selected nfxC-type mutants). R1–R7 are wild-typeresistant isolates.
Table 5. Comparison of mRNA concentrations of nfxC-like
are regulated individually to some extent. The levels of
mRNA of these genes may, therefore, not be exactly in accordwith protein levels. However, the quantification results with
RT-PCR in this study were in agreement with the expected
expression profile of nfxC-type mutants.
Resistance due to the altered regulation of intrinsic genes in
P. aeruginosa is not well understood. This type of resistance
may depend on the regulation of a more composite co-
regulated network of multiple operons as well as the
‘quorum-sensing systems’ of P. aeruginosa
2001ortunately, our understanding of this
is extremely limited due to the limited power of the methods
used at present. Therefore, further studies using new meth-
ods deserve increased scientific interest. We believe that real-
time quantification with the selection of suitable internal
control genes will facilitate studies and provide new insights
into the regulatory alterations of innate genes and themultiple antibiotic resistance problem of P. aeruginosa.
This study showed that proC and rpoD form the most stablepair in a set of clonally unrelated P. aeruginosa strains with
protein. It is known that mRNA expression predicts mature
diverse resistance phenotypes. Thus, this pair may be used as
protein levels poorly and there may be up to 30-fold
internal controls in relative comparison studies of resistance
differences ). Transcription and translation
Negative regulation of the Pseudomonas aeruginosa outer membraneporin OprD selective for imipenem and basic amino acids. Antimicrob
We are grateful to Dr Serhat Unal (Hacettepe Universitesi), Dr Mehmet
Ali Inal (Ege Universitesi) and Dr Hakan Leblebicioglu (Ondokuz MayısUniversitesi) for providing strains and to Jason John Nash and Victor L.
Okamoto, K., Gotoh, N. & Nishino, T. (2001). Pseudomonas aeruginosa
Yu for English reading of this manuscript.
reveals high intrinsic resistance to penem antibiotics: penem resistancemechanisms and their interplay. Antimicrob Agents Chemother 45,1964–1971.
Poole, K. (2001). Multidrug efflux pumps and antimicrobial resistance
Gygi, S. P., Rochon, Y., Franza, B. R. & Aebersold, R. (1999).
in Pseudomonas aeruginosa and related organisms. J Mol Microbiol
Correlation between protein and mRNA abundance in yeast. Mol Cell
Renders, N., Romling, Y., Verbrugh, H. & van Belkum, A. (1996).
Jacobs, C., Joris, B., Jamin, M. & 7 other authors (1995). AmpD,
Comparative typing of Pseudomonas aeruginosa by random amplifica-
essential for both beta-lactamase regulation and cell wall recycling, is a
tion of polymorphic DNA or pulsed-field gel electrophoresis of DNA
novel cytosolic N-acetylmuramyl-L-alanine amidase. Mol Microbiol 15,
macrorestriction fragments. J Clin Microbiol 34, 3190–3195.
Schnider, U., Keel, C., Blumer, C., Troxler, J., Defago, G. & Haas, D.
Johnson, M. R., Wang, K., Smith, J. B., Heslin, M. J. & Diasio, R. B.
(1995). Amplification of the housekeeping sigma factor in Pseudomonas
(2000). Quantitation of dihydropyrimidine dehydrogenase expression
fluorescens CHA0 enhances antibiotic production and improves bio-
by real-time reverse transcription polymerase chain reaction. Anal
control abilities. J Bacteriol 177, 5387–5392.
Kerr, J. R., Moore, J. E., Curran, M. D., Graham, R., Webb, C. H., Lowry, K.
Sumita, Y. & Fukasawa, M. (1996). Meropenem resistance in Pseudo-
G., Murphy, P. G., Wilson, T. S. & Ferguson, W. P. (1995). Investigation
monas aeruginosa. Chemotherapy 42, 47–56.
of a nosocomial outbreak of Pseudomonas aeruginosa pneumonia in an
Trias, J., Dufresne, J., Levesque, R. C. & Nikaido, H. (1989). Decreased
intensive care unit by random amplification of polymorphic DNA assay.
outer membrane permeability in imipenem-resistant mutants of
Pseudomonas aeruginosa. Antimicrob Agents Chemother 33, 1202–1206.
Kohler, T., van Delden, C., Curty, L. K., Hamzehpour, M. M. & Pechere, J.
Vahaboglu, H., Ozturk, R., Akbal, H., Saribas, S., Tansel, O. &
C. (2001). Overexpression of the MexEF-OprN multidrug efflux system
Coskunkan, F. (1998). Practical approach for detection and identifica-
affects cell-to-cell signaling in Pseudomonas aeruginosa. J Bacteriol 183,
tion of OXA-10-derived ceftazidime-hydrolyzing extended-spectrum
beta-lactamases. J Clin Microbiol 36, 827–829.
Kutchma, A. J., Hoang, T. T. & Schweizer, H. P. (1999). Characterization
Vandecasteele, S. J., Peetermans, W. E., Merckx, R. & Van Eldere, J.
of a Pseudomonas aeruginosa fatty acid biosynthetic gene cluster:
(2001). Quantification of expression of Staphylococcus epidermidis
purification of acyl carrier protein (ACP) and malonyl-coenzyme
housekeeping genes with Taqman quantitative PCR during in vitro
A : ACP transacylase (FabD). J Bacteriol 181, 5498–5504.
growth and under different conditions. J Bacteriol 183, 7094–7101.
Livermore, D. M. (1992). Interplay of impermeability and chromosomalbeta-lactamase activity in imipenem-resistant Pseudomonas aeruginosa.
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De
Antimicrob Agents Chemother 36, 2046–2048.
Paepe, A. & Speleman, F. (2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple
Maseda, H., Yoneyama, H. & Nakae, T. (2000). Assignment of the
internal control genes. Genome Biol 3, research0034.1-0034.11. http://
substrate-selective subunits of the MexEF-OprN multidrug efflux pump
of Pseudomonas aeruginosa. Antimicrob Agents Chemother 44, 658–664.
Masuda, N., Sakagawa, E., Ohya, S., Gotoh, N., Tsujimoto, H. &
Nishino, T. (2000). Substrate specificities of MexAB-OprM, MexCD-
TaqMan polymerase chain reaction: validation and comparison with
OprJ, and MexXY-oprM efflux pumps in Pseudomonas aeruginosa.
RNase protection. Anal Biochem 269, 198–201.
Antimicrob Agents Chemother 44, 3322–3327.
Wei, Y., Lee, J. M., Richmond, C., Blattner, F. R., Rafalski, J. A. &
Morita, Y., Komori, Y., Mima, T., Kuroda, T., Mizushima, T. & Tsuchiya,
LaRossa, R. A. (2001). High-density microarray-mediated gene ex-
T. (2001). Construction of a series of mutants lacking all of the four
pression profiling of Escherichia coli. J Bacteriol 183, 545–556.
major mex operons for multidrug efflux pumps or possessing each one
Yucesoy, M., Yulug, N., Kocagoz, S., Unal, S., Cetin, S. & Calangu, S.
of the operons from Pseudomonas aeruginosa PAO1: MexCD-OprJ is an
(2000). Antimicrobial resistance of gram-negative isolates from
inducible pump. FEMS Microbiol Lett 202, 139–143.
intensive care units in Turkey: comparison to previous three years.
Ochs, M. M., McCusker, M. P., Bains, M. & Hancock, R. E. (1999).
VOLUNTARY STOPPING EATING & DRINKING This paper addresses questions about stopping eating and drinking as an option to hasten death. The U.S. Supreme Court has affirmed the right of a competent individual to refuse medical therapies and this includes food and fluids. This choice is also commonly accepted in the medical community. One of the advantages of this decision is that you may chan
Kapitel 91, Jeg var klar til at leve mit liv endnu gang: Forandret mig meget havde jeg ikke. Stadig uden trusser og med hul i nederdelens lommer. Fem fingre i kussen. Efter at min ven var død, kvast til hakkemad i en trafikulykke i begyndelsen af juni 1998 og min datters død 2000 gik der mange år før jeg igen var klar til at udfordre livet. På kalenderen stod der nu 2007

 Housekeeping genes of Pseudomonas aeruginosa
Table 1. Primers used in quantification studies
calculation of Spearman’s correlation coefficients of the raw concentra-tion data with the aid of the statistical package SPSS (version 9.0). The
best-correlated pair was considered to be the most stable one.
Housekeeping genes of Pseudomonas aeruginosa
Table 1. Primers used in quantification studies
calculation of Spearman’s correlation coefficients of the raw concentra-tion data with the aid of the statistical package SPSS (version 9.0). The
best-correlated pair was considered to be the most stable one.