Le métronidazole (Flagyl) reste la référence dans le traitement des infections anaérobies et des parasitoses comme la giardiase ou l’amibiase. Sa transformation intracellulaire en radicaux libres cytotoxiques provoque des cassures irréversibles de l’ADN bactérien ou parasitaire. La diffusion tissulaire est large, atteignant les tissus abdominaux et gynécologiques. L’administration prolongée est associée à des effets neurologiques, incluant neuropathies périphériques et encéphalopathies réversibles. L’association avec l’alcool déclenche une réaction de type antabuse. Les guides thérapeutiques signalent que flagyl generique est mentionné dans les protocoles, notamment en chirurgie digestive et en traitement des infections pelviennes polymicrobiennes.
037155u716
Copyright 2004 by the Genetics Society of AmericaDOI: 10.1534/genetics.104.026716
Virulence, Multiple Infections and Regulation of Symbiotic Population in the Wolbachia-Asobara tabida Symbiosis Laurence Mouton,*,1 Franck Dedeine,*,† He´le`ne Henri,* Michel Boule´treau,* Ne´rina Profizi‡ and Fabrice Vavre*
*Laboratoire de Biome´trie et Biologie Evolutive (UMR-CNRS 5558), Universite´ Claude Bernard—Lyon1, 69622 Villeurbanne Cedex, France,
†Department of Entomology, University of Wisconsin, Madison, Wisconsin 53706 and
‡CHI de Toulon-La Seyne sur Mer, 83507 La Seyne sur Mer, France
The density and regulation of microbial populations are important factors in the success of symbiotic
associations. High bacterial density may improve transmission to the next generation, but excessive replica-tion could turn out to be costly to the host and result in higher virulence. Moreover, differences invirulence may also depend on the diversity of symbionts. Using the maternally transmitted symbiontWolbachia, we investigated how bacterial density and diversity are regulated and influence virulence inhost insects subject to multiple infection. The model we used was the wasp Asobara tabida that naturallyharbors three different Wolbachia strains, of which two are facultative and induce cytoplasmic incompatibil-ity, whereas the third is necessary for the host to achieve oogenesis. Using insect lines infected withdifferent subsets of Wolbachia strains, we show that: (i) some traits of A. tabida are negatively affected byWolbachia; (ii) the physiological cost increases with the number of co-infecting strains, which also corre-sponds to an increase in the total bacterial density; and (iii) the densities of the two facultative Wolbachiastrains are independent of one another, whereas the obligatory strain is less abundant when it is alone,suggesting that there is some positive interaction with the other strains.
THE genetic diversity of parasites co-infecting indi- reasonsforthis.First,theverticaltransmissionofsymbi-
vidual hosts is often thought to be an important
onts tends to limit multiple infection, because the oo-
factor in the evolution of their virulence (Ewald 1994;
cytes of the host are colonized by only a few individuals,
Frank 1996a; Galvani 2003). Theoretical studies have
and the resulting bottleneck greatly reduces the genetic
shown that lower relatedness among the parasites within
diversity of the symbiotic population (Mira and Moran
the host could lead to increased virulence (Frank 1994,
2002). Second, many vertically transmitted symbionts
1996a; Van Baalen and Sabelis 1995). The classical
have evolved a mutualistic relationship with their hosts,
idea, referred to as the “tragedy of the commons,” is
and the benefit they confer considerably outweighs their
that competition for a limiting resource puts faster ex-
cost; this makes the cost difficult to assess or even detect
ploiters at an advantage over more prudent ones (Har-
(Thompson 1988; Bronstein 1994). However, these
din 1968). However, recent articles have pointed out
biological models could provide interesting data, be-
that in commonly used models, selection among para-
cause the closely linked evolutionary fates of host and
sites affects only the host exploitation rate, but that
symbionts should have led to the selection of mecha-
different outcomes could be reached when other types
nisms that reduce virulence to a minimum (Lipsitch
of competition are also considered (Chao et al. 2000;
Read and Taylor 2001; Brown et al. 2002). For exam-
A good model for studying this question is the mater-
ple, interference among parasites could lead to under-
nally transmitted symbiotic bacterium Wolbachia, which
exploitation of hosts and hence to reduced virulence
is able to induce cytoplasmic incompatibility (CI), lead-
(Chao et al. 2000). The relationship beween virulence
ing to postzygotic reproductive isolation between any
and multiple infection is thus still under debate and
male infected by a Wolbachia strain and a female lacking
needs further empirical documentation.
this strain (for review see Hoffmann and Turelli 1997).
So far these questions have received little study in
This puts females with multiple infection at an advan-
vertically transmitted symbionts. There are two main
tage and promotes the spread and maintenance of mul-tiple infection (Frank 1998), which has proved rathercommon. Second, infected individuals benefit only in-
1Corresponding author: Laboratoire de Biome´trie et Biologie Evolutive
directly from infection, as a result of the disadvantage
(UMR-CNRS 5558), Universite´ Claude Bernard—Lyon1 43, Bd. du
suffered by uninfected females or females infected by
11 Novembre 1918, 69622 Villeurbanne Cedex, France. E-mail: mouton@biomserv.univ-lyon1.fr
only a subset of Wolbachia strains. Thus, the cost of
Genetics 168: 181–189 (September 2004)
and so most of their oocytes are already mature at emergence.
infection (that we define here as the virulence of Wol-
To estimate the oocyte load, newly emerged females were kept
bachia) and the advantage to the host are clearly distin-
for 5 days with water and honey to allow oocyte maturation
guishable and can be measured separately in distinct
to be completed. The ovaries were dissected in physiological
saline, transferred into neutral red solution for 5 min, and
In this study, we used the association between Wol-
then gently squashed between the slide and cover glass todisperse the eggs. The stained eggs were then counted under
bachia and the parasitic wasp Asobara tabida, whose lar-
the microscope with the help of a video system.
vae develop as solitary endoparasites of Drosophila lar-
Tibia length: Left hind tibia was measured on the adult males
vae. A. tabida individuals are co-infected by three
and females of each line using a micrometer.
different Wolbachia strains, wAtab1, wAtab2, and wAtab3Dry weight: Emerging parasitoids of both sexes were sampled
(Vavre et al. 1999). While both wAtab1 and wAtab2 are
in each line and dried at 65Њ for 48 hr before weighing. Adult ability to survive starvation: Two-day-old males and fe-
facultative parasites and induce CI (F. Dedeine, per-
males were put in vials with moist cotton, but without food.
sonal communication), wAtab3 is necessary for oogen-
Mortality was checked every day at the same time (Ϯ1 hr)
esis to occur in this species: A. tabida females lacking
until all the individuals had died. Six vials, each containing
wAtab3 do not produce oocytes (Dedeine et al. 2001).
10 parasitoids, were studied for each sex in each line.
Using antibiotherapy, we created A. tabida lines that
Locomotor activity pattern: Individual locomotor activity was
monitored using a video-tracking and image-analysis system
harbored different combinations of Wolbachia strains,
to provide automatic continuous measurements of the insects
but shared the same nuclear background.
over several days (Allemand et al. 1994). Individual insects
This study addresses two main questions: (i) Is there
were isolated in experimental circular glass arenas with honey
any relationship between the diversity and virulence
as food. The locomotor activity of each individual was mea-
of Wolbachia and/or between Wolbachia density and
sured every 3 min using binary data (1 if the wasp had movedduring a 2-sec video recording and 0 if it had not), and the
virulence? and (ii) How are Wolbachia diversity and
hourly activity was calculated as the percentage of active re-
density regulated? The findings are discussed in the
cordings obtained over 3 days with a 12/12 hr LD cycle. To
context of the evolution of virulence in multiple infec-
evaluate the average daily pattern of activity, two independent
tions, by considering the peculiar selective pressures
parameters were estimated for each individual: the rate of
locomotor activity, calculated as the average of active re-cordings over a 24-hr period, and the profile of the rhythm,which establishes the pattern of the total daily activity in termsof the hourly percentages. The rate of locomotor activity mea-
sures the locomotor performance of wasps, while the activityprofile determines how this activity is organized throughout
Insect strains and rearing: A. tabida (Hymenoptera: Braco-
the day. Data are reported in terms of the Zeitgeber time (Zt,
nidae) is a solitary endoparasitoid wasp of several Drosophila
time within the environmental cycle); the light is turned off
species. In the laboratory, parasitoids are reared on a Wol-
bachia-free strain of Drosophila melanogaster originating from
Real-time quantitative PCR: DNA extraction: Insects or parts
Ste Foy-les-Lyon (France). Rearing and experiments were car-
of insects were individually squashed in 150 l 5% (w/vol)
ried out without larval or adult competition at 20Њ under 12/12 light/dark (LD) cycle and 70% relative humidity.
Chelex solution (Bio-Rad, Richmond, CA) and proteinase K
A. tabida individuals are naturally infected with three Wol-
(Eurobio, Les Ulis, France; final concentration 0.5 g/l)
bachia strains, named wAtab1, wAtab2, and wAtab3 (Vavre et
and kept at 56Њ for 6 hr. After 15 min at 95Њ, the samples were
al. 1999). The triply infected line, named Pi(123), is an inbred
centrifuged at 16,000 ϫ g for 4 min.
line originating from Pierrefeu (Pi), France, which has been
Primers: Quantification of Wolbachia bacteria was achieved
maintained by regular sib-mating for four generations. De-
by amplifying the Wolbachia surface protein gene wsp. To
rived A. tabida lines with different infection statuses were
detect all the Wolbachia strains present we used the general
obtained using moderate antibiotic treatments (F. Dedeine,
forward primers 81F: 5Ј-TGG TCC AAT AAG TGA TGA AGA
personal communication). Since wAtab3 is obligatory for re-
AAC-3Ј (Braig et al. 1998). Specific PCR detection of each
production in this species, only one singly infected and two
Wolbachia strain was conducted using three other forward
doubly infected lines could be obtained and have proved stable
primers: 165ЈF (5Ј-TGG TAT TAC AAA TGT AGC-3Ј) for wA-
in time: the singly infected line Pi(3) harboring wAtab3 and
tab1, 172F (5Ј-ACC TAT AAG AAA GAC AAG-3Ј) for wAtab2
the doubly infected lines Pi(13) and Pi(23) harboring wAtab3
(Zhou et al. 1998), and Aso3 (5Ј-AAA GGG GAC TGA TGA
and wAtab1 and wAtab3 and wAtab2, respectively.
TGT-3Ј) for wAtab3. All these forward primers were used with
Components of the fitness cost of infection: Several fitness
the same reverse primer 691R: 5Ј-AAA AAT TAA ACG CTA
traits have been measured in individuals of each line of A.
CTC CA-3Ј (Zhou et al. 1998). Quantitative PCR: Real-time quantitative PCR was performed
Offspring production and sex ratio: After mating with 3- to
using the LightCycler system (Roche). The 20-l reaction mix-
4-day-old males with the same infection status as themselves
ture consisted of 10% (vol/vol) LightCycler DNA master SYBR
(checked by individual visual inspection), 1- to 2-day-old fe-
Green I (Roche Diagnostics), 3 mm MgCl2, 500 nm each
males were each provided with 150 Drosophila larvae (24 hr
primer, and 2 l of template DNA. The amplification consisted
old) and allowed to parasitize hosts for 48 hr. The infested
of 40 cycles of 15 sec at 95Њ, followed by 14 sec at 53Њ and 28
host larvae were then kept and allowed to develop. At emer-
sec at 72Њ for 81F/691R and 11 sec at 52Њ and 22 sec at 72Њ
gence, the adult Drosophila and the wasps of both sexes were
for 165ЈF/691R, 172F/691R, and Aso3/691R.
counted in each vial. We performed three series (i.e., blocks)
Standard curves were drawn on clones of the three Wol-
to test these traits (at least seven replicates per line for each
bachia strains. Amplification with 81F/691R primers was per-
formed on a triply infected female. PCR products were puri-
Egg production: A. tabida females are mainly proovogenic,
fied (GIBCO BRL, Gaithersburg, MD) and cloned into the
Virulence, Multiple Infections and Regulation of Symbiotic Population
Pi(123),
Figure 1.—Starvation survival curves of female and male
Figure 2.—Mean curves of the locomotor activity rhythms
wasps with differing Wolbachia infection statuses. (ᮀ) Pi(123),
of A. tabida lines Pi(123), Pi(13), Pi(23), and Pi(3). Males and
(᭡) Pi(13), (᭜) Pi(23), (᭺) Pi(3).
females were measured for 3 days under LD 12/12 (Zeitgebertime) with light off at Zt0 and light on at Zt12. The black-and-white rectangles at the top of the figure represent night
pDrive cloning vector (QIAGEN, Valencia, CA). Specific PCR
and day, respectively. (ᮀ) Pi(123), (᭡) Pi(13), (᭜) Pi(23),
assays were used to identify the Wolbachia strain present in
the clone. The DNA concentration of each sample was mea-sured by OD absorbance at 260 nm. Standard curves wereplotted using five dilutions of this vector (from 102 to 108
and Pi(123) were the lightest. Thus, the greater the
copies) containing one copy of a specific wsp sequence, which
diversity of Wolbachia lineages harbored by the insects,
is a single-copy gene (Braig et al. 1998). The number of
the less they weighed. More diversity in Wolbachia also
Wolbachia cells was calculated as described in Noda et al. (2001). These values must be considered as semiquantitative
led to a shorter life span of the host (Table 1; Figure
estimates of Wolbachia cell numbers.
1). Life span of doubly infected wasps was intermediate
Wolbachia abundance was measured individually on 5-day-
for both sexes, but the difference from simply infected
old males and females, either in whole bodies or separately
wasps was not significant in the males. The rate of loco-
in head plus thorax and abdomen of the same individual.
motor activity also varied with the infection status of
Density was obtained by correcting the number of Wolbachiaby the mean fresh weight of insects of the line. The ratio of
individuals (Table 1). As for dry weight and life span,
Wolbachia cells in the abdomen to the sum of Wolbachia in
singly infected wasps had greater locomotor activity than
head plus thorax and abdomen was calculated individually.
triply infected ones, but in this case the difference wasnot significant for the females. In contrast, the profileof the rhythm was the same regardless of infection status
(ANOVA, d.f. ϭ 3189, F ϭ 1.763, P ϭ 0.16, Figure 2). Infection cost: Among the seven fitness components Total Wolbachia density: Previous findings have
studied here, three do not vary according to the infec-
shown that infection cost increases with bacterial diver-
tion status: productivity, fecundity, and tibia length. For
sity. To find out whether this is linked to density varia-
sex ratio, a marginally significant effect was detected,
tions, we measured the number of Wolbachia cells in
with a potentially higher proportion of females in the
male and female Pi(3), Pi(13), Pi(23), and Pi(123) (Ta-
triply infected line. However, this trait is highly variable
ble 2). In all these lines, the numbers of Wolbachia and
and no such effect was detected in the third block. Clear
their relative densities (number of cells per milligram
conclusions were obtained only for dry weight, adult
of fresh weight) were higher in females than in males
survival, and rate of locomotor activity (Table 1).
(Mann-Whitney test, P Ͻ 0.001).
Pi(3) individuals of both sexes were the heaviest,
Despite the fact that the cell numbers obtained by
Pi(13) and Pi(23) had similar and intermediate weights,
real-time quantitative PCR were rather low and may have
Virulence, Multiple Infections and Regulation of Symbiotic Population
Wolbachia density in A. tabida species
Mean values ϮSE and statistical analysis of the total number of Wolbachia cells per wasp (eight replicates
per sex and line) are shown. The analyses were carried out using the Kruskal-Wallis nonparametric test. Meansmarked with the same letter are not significantly different (Mann-Whitney tests, P ϭ 0.05).
been underestimated, in both sexes the total Wolbachia
there was no significant difference between males of
density depended on the combination of the Wolbachia
the two doubly and the one triply infected lines.
strains co-infecting the same individual host (Kruskal-
Overall, these findings demonstrate that increasing
Wallis test, P Ͻ 0.005). Singly infected individuals of
the mixture of Wolbachia strains results in a higher
the Pi(3) strain had the lowest density, which was less
physiological cost to the host and also leads to higher
than one-quarter of that in Pi(123). In females, the
Wolbachia density. This means that the cost of infection
density in the two doubly infected lines was intermediate
is also positively correlated with bacterial density, as
between those of the singly and triply infected lines,
but it was lower in Pi(13) than in Pi(23). In contrast,
Strain-specific Wolbachia density: We have shown that
Figure 3.—Wolbachia density and dry weight.
The Wolbachia density (histogram) and dryweight (curve) of Pi(123), Pi(13), Pi(23), andPi(3) individuals are presented. Values corre-spond to the average of eight individuals for den-sity and of 30 individuals for dry weight. Bars showthe standard error.
Figure 4.—Specific densities of wAtab1, wAtab2, and wAtab3. Specific densities of wAtab1, wAtab2, and wAtab3 in females and
males of various infection statuses are presented. Values correspond to the average of eight individuals per sex and line. Barsshow the standard error. P-values of Kruskal-Wallis tests are indicated. For wAtab3, means marked with the same letters are notsignificantly different (Mann-Whitney tests, P ϭ 0.05).
total bacterial density increases with bacterial diversity,
Considering the relative abundance of Wolbachia
but we still do not know how each Wolbachia strain
strains in triply infected individuals, wAtab1 was found
responds to the presence of other Wolbachia strains.
to be the least represented in both sexes (16% in females
We therefore measured the specific density of each Wol-
and 22% in males), whereas the most abundant strains
bachia strain co-infecting the same individual in all A.
were wAtab2 in females (61%) and wAtab3 in males
(near 41%). Despite the different abundances of wAtab1
First, the sums of the specific densities are equal to
and wAtab2 in the host, they induced similar infection
the estimations of total densities in all lines (Wilcoxon
rank test, P ϭ 0.11), thus indicating the reliability of
Wolbachia distribution in the host body: The localiza-
the method and ruling out any concern about signifi-
tion of Wolbachia may have an influence on the infec-
tion cost (McGraw et al. 2002). We then studied the
In both sexes, we found that the specific densities of
preferential localization of these bacteria in A. tabidawAtab1 and wAtab2 were the same in the Pi(123) and
species of each line by measuring the total number of
Pi(13) lines and the Pi(123) and Pi(23) lines (Mann-
Wolbachia cells in head plus thorax and in abdomen
Whitney test, P Ͼ 0.05), respectively, and are thus inde-
of the same host body. Comparison of the percentage
pendent of the infection status of the individuals. The
of Wolbachia in abdomen compared to the entire body
density of wAtab3 was lower in Pi(3) than in multiply
in the four lines demonstrates that Wolbachia are pref-
infected lines and, whereas the differences were not
erentially localized in abdomen in males as well as in
significant in the males (Kruskal-Wallis test, P ϭ 0.11),
females whatever the infection status (Table 3). How-
they were significant in the females (P ϭ 0.02), with the
ever, the percentage of Wolbachia in abdomen is higher
Pi(13) and Pi(123) lines harboring more wAtab3 than
in females than in males (Student’s t-test, P Ͻ 0.0001).
Pi(3) lines (Mann-Whitney test, P Յ 0.02).
This repartition does not differ between lines (ANOVA,
Virulence, Multiple Infections and Regulation of Symbiotic Population
though variations in density are due to the differingdiversity of the Wolbachia strains infecting individu-
Percentage of Wolbachia cells in abdomen compared to the entire body in A. tabida females and males
als, higher bacterial densities also correlate with anincrease in the cost of infection. A relationship be-
tween density and virulence has already been docu-
mented by McGraw et al. (2002), who demonstratedthat the fitness cost associated with the popcorn strain
was reduced in a new transfected Drosophila host
harboring a lower Wolbachia density. Therefore,
with results of McGraw et al. (2002), the data pre-
sented here strongly support the existence of a rela-
tionship between bacterial density and infection cost,
which has rarely been demonstrated hitherto, but
is generally accepted since more symbionts can be
expected to require more energy (Thompson 1988). However other factors, such as the particular traits
Mean percentage ϮSE and statistical analysis of total Wol-
of each strain, may affect the impact of bacteria on
bachia cells in abdomen compared to the sum of densities inhead (H), thorax (T), and abdomen (A) in five males and
their host. For example, the same cost is induced by
five females of each A. tabida line are shown. One-way ANOVA
wAtab1 and wAtab2, in spite of the lower density of
analyses were carried out after arcsine square root transforma-
wAtab1. Thus, all differences in the cost of infection
cannot be attributed to bacterial density alone.
iii. The densities of the two facultative strains wAtab1P Ͼ 0.05 for both sexes), even though the percentage
and wAtab2 are specifically regulated and do not
of Wolbachia in abdomen of males for the Pi(3) line
depend on the presence of other strains. The situa-
tion is less clear for the obligatory bacterium wAtab3,suggesting that there may be some positive interac-tion with other strains. Such specific bacterial regu-
lation seems to be rather common in multiple infec-
Three main conclusions can be drawn from our find-
tions by Wolbachia, as suggested by studies in D.simulans, Ephestia kuehniella, and Leptopilina hetero-toma (Rousset et al. 1999; Ikeda et al. 2003; Mouton
i. Despite the vertical transmission of Wolbachia and
the high selective pressures that tend to reduce infec-tion costs, some traits of A. tabida are still negatively
The puzzling question that now arises is how the infec-
affected by Wolbachia. However, the resulting costs
tion cost for A. tabida has persisted despite selective
are low, may have little influence on the fitness of
pressures toward reduced virulence that act on both the
adults in the wild, and probably do not affect the
bacteria and the insects. Is multiple infection responsi-
maintenance of multiple infection. This finding is
ble for this persistence? The strain-specific regulation
consistent with numerous studies on the cost of Wol-
of bacterial density suggests that strains do not compete
bachia where a high variability among species and
with one another for limited resources within the host,
traits exists (Hoffmann et al. 1990, 1994, 1998; Gior-
and this should prevent the increased virulence that is
dano et al. 1995; Girin and Boule´treau 1995; Tur-
usually expected from multiple infection (Frank 1996a).
elli and Hoffmann 1995; Wade and Chang 1995;
The maintenance of infection cost in A. tabida probably
Bourtzis et al. 1996; Clancy and Hoffmann 1997;
expresses specific constraints, such as the classical trade-
Min and Benzer 1997; Poinsot and Merc¸ot 1997;
off between virulence and bacterial transmission (for
Hoerauf et al. 1999; Vavre et al. 1999; Fleury et al.
review, see Frank 1996b). On the one hand, bacterial
2000; Dobson et al. 2002; Fry and Rand 2002). On
density should be kept as low as possible to reduce
the other hand, the nearly significant effect on sex
fitness costs to the host, but on the other hand, the
ratio might promote multiple infections through in-
intracellular density of microorganisms must be high
crease in the proportion of females.
enough to ensure transmission to the next generation.
ii. The cost of infection depends directly on the diver-
sity of Wolbachia and increases with the number of
Moreover, convergent selective pressures also act on the
Wolbachia strains within the insect host. The same
host (Turelli 1994; Vavre et al. 2003), since strain loss
localization has been observed between individuals
can have dramatic indirect effects on individual fitness.
of all infection statuses; therefore the differences of
In A. tabida, this is obvious for the obligatory wAtab3
infection cost observed in the various lines of A.
strain, since loss of this strain results in female sterility. tabida cannot be explained by a difference of localiza-
It is also true to some extent for the two facultative
tion in the host body. These results mean that even
strains, both of which induce high CI levels (Ͼ70%; F.
Fleury, F., F. Vavre, N. Ris, P. Fouillet and M. Boule´treau, 2000
Dedeine, personal communication). Losing one or
Physiological cost induced by the maternally-transmitted endo-
both of them can be expected to expose females to CI
symbiont Wolbachia in the Drosophila parasitoid Leptopilina heter-
and counterselection, whereas it should have no effect
otoma. Parasitology 121: 493–500.
Recognition and polymorphism in host-parasite
on males. Consequently, selective pressures do promote
genetics. Philos. Trans. R. Soc. Lond. B Biol. Sci. 346: 283–293.
the maintenance of diversity of infection, and this could
Host-symbiont conflict over the mixing of symbi-
explain how the specificity of density regulation has
otic lineage. Proc. R. Soc. Lond. B Biol. Sci. 263: 339–344.
Models of parasite virulence. Q. Rev. Biol. 71:
evolved in spite of other types of competition, such as
interference between Wolbachia strains. In contrast, the
Dynamics of cytoplasmic incompatibility with
density data for wAtab3 suggest that there may be some
multiple Wolbachia infections. J. Theor. Biol. 192: 213–218.
positive interaction between this obligatory strain and
mine Drosophila melanogaster survival. Evolution 56: 1976–1981.
the two facultative ones, since the density of wAtab3 is
Epidemiology meets evolutionary ecology.
lower when it is the only strain present.
Trends Ecol. Evol. 18: 132–139.
Giordano, R., S. L. O’Neill and H. M. Robertson, 1995
Finally, the bacterial density regulating system exhib-
infections and the expression of cytoplasmic incompatibility in
ited by A. tabida and other multiply infected species may
Drosophila sechellia and D. mauritiana. Genetics 140: 1307–1317.
limit both competition for resources and interference
ation in host infestation efficiency in a parasitoid wasp Tricho-
between different Wolbachia strains, and this may re-
gramma bourarachae. Experientia 52: 398–402.
flect the peculiar selective pressures acting on the sys-
The tragedy of the commons: the population
problem has no technical solution; it requires a fundamental
tem. Multiple infection by Wolbachia can be viewed as
extension in morality. Science 162: 1243–1248.
a criminal conspiracy among bacteria, with each partner
Hoerauf, A., K. Nissen-Pa¨hle, C. Schmetz, K. Henkle-Du
relying on the others for its own fitness. Clearly, the
lular bacteria in the filarial nematode Litomosoides sigmodontis and
success of the plot relies on the reciprocal agreement
results in filarial infertility. J. Clin. Invest. 103: 11–17.
among accomplices, and reciprocal damage would be
in insects, pp. 42–80 in Influential Passengers: Inherited Microorgan-isms and Arthropod Reproduction, edited by S. L. O’Neill, A. A.
We thank the De´veloppement Technologique et Analyse Mole´cu-
Hoffmann and J. H. Werren. Oxford University Press, Oxford.
laire de la Biodiversite´ for use of the Light Cycler System. This study
Hoffmann, A. A., M. Turelli and L. G. Harshman, 1990
was partly supported by Centre National de la Recherche Scientifique
affecting the distribution of cytoplasmic incompatibility in Dro-sophila simulans. Genetics 126: 933–948.
Hoffmann, A. A., D. Clancy and E. Merton, 1994
incompatibility in Australian populations of Drosophila melanogas- ter. Genetics 136: 993–999.
Hoffmann, A. A., M. Hercus and H. Dagher, 1998
dynamics of the Wolbachia infection causing cytoplasmic incom- patibility in Drosophila melanogaster. Genetics 148: 221–231.
Allemand, R., F. Pompanon, F. Fleury, P. Fouillet and M. Bou-
Ikeda, T., H. Ishikawa and T. Sasaki, 2003
Behavioural circadian rhythms measured in real-
density in the Mediterranean flour moth, Ephestia kuehniella, and
time by automatic image analysis: application in parasitoid insects.
the almond moth, Cadra cautella. Zool. Sci. 20: 153–157.
Parasitol. Entomol. 19: 1–8.
Lipsitch, M., S. Siller and M. A. Nowak, 1995
Bourtzis, K., A. Nirgianaki, G. Markakis and C. Savakis, 1996
virulence in pathogens with vertical and horizontal transmission.
Wolbachia infection and cytoplasmic incompatibility in Drosoph-
Evolution 50: 1729–1741.
ila species. Genetics 144: 1063–1073.
McGraw, E. A., D. J. Merritt, J. N. Droller and S. L. O’Neill,
Bourtzis, K., H. R. Braig and T. L. Karr, 2003
Wolbachia density and virulence attenuation after transfer
patibility, pp. 217–247 in Insect Symbiosis, edited by T. Miller and
into a novel host. Proc. Natl. Acad. Sci. USA 99: 2918–2923.
K. Bourtzis. CRC Press, Boca Raton, FL.
Braig, H. R., W. Zhou, S. L. Dobson and S. L. O’Neill, 1998
of Drosophila, can be virulent, causing degeneration and early
ing and characterization of a gene encoding the major surface
death. Proc. Natl. Acad. Sci. USA 94: 10792–10796.
protein of the bacterial endosymbiont Wolbachia pipientis. J. Bacte-
riol. 180: 2373–2378.
transmission bottlenecks in maternally transmitted endosymbio-
Conditional outcomes in mutualistic interac-
tic bacteria. Microb. Ecol. 44: 137–143.
tions. Trends Ecol. Evol. 9: 214–217.
Mouton, L., H. Henri, M. Boule´treau and F. Vavre, 2003
Brown, S. P., M. E. Hochberg and B. T. Grenfell, 2002
specific regulation of intracellular Wolbachia density in multiply
multiple infection select for raised virulence? Trends Microbiol.
infected insects. Mol. Ecol. 12: 3459–3465. 10: 401–405.
Noda, H., Y. Koizumi, Q. Zhang and K. Deng, 2001
Chao, L., K. A. Hanley, C. L. Burch, C. Dahlberg and P. E. Turner,
sity of Wolbachia and incompatibility level in two planthopper
Kin selection and parasite evolution: higher and lower
species, Laodelphax striatellus and Sagatella furcifera. Insect Bio-
virulence with hard and soft selection. Q. Rev. Biol. 75: 261–275.
chem. Mol. Biol. 31: 727–737.
endosymbionts from Drosophila simulans in Drosophila serrata, a
simulans: Does the female host bear a physiological cost? Evolu-
novel host. Am. Nat. 149: 975–988.
tion 51: 180–186.
Dedeine, F., F. Vavre, F. Fleury, B. Loppin, M. E. Hochberg et
Removing symbiotic Wolbachia bacteria specifically
diverse infections. Science 292: 1099–1102.
inhibits oogenesis in a parasitic wasp. Proc. Natl. Acad. Sci. USA
Rousset, F., H. R. Braig and S. L. O’Neill, 1999
98: 6247–6252.
Wolbachia infection in Drosophila with nearly additive incompati-
Dobson, S. L., E. J. Marsland and W. Rattanadechakul, 2002
bility effects. Heredity 82: 620–627.
Mutualistic Wolbachia infection in Aedes albopictus : accelerating
Variation in interspecific interactions. Annu.
cytoplasmic drive. Genetics 160: 1087–1094.
Rev. Ecol. Syst. 19: 65–87. Evolution of Infectious Disease. Oxford University
Evolution of incompatibility-inducing microbes
and their hosts. Evolution 48: 1500–1513.
Virulence, Multiple Infections and Regulation of Symbiotic Population
host species selection on cytoplasmic incompatibility-inducing
in Drosophila simulans: dynamics and parameter estimates from
Wolbachia in haplodiploids. Evolution 57: 421–427.
natural populations. Genetics 140: 1319–1338. Tribolium confusum beetles after infection with the intracellular
infection and the evolution of virulence. Am. Nat. 146: 881–910.
parasite Wolbachia. Nature 373: 72–74.
Vavre, F., F. Fleury, D. Lepetit, P. Fouillet and M. Boule´treau,
Zhou, W., F. Rousset and S. O’Neill, 1998
Phylogenetic evidence for horizontal transmission of Wol-
based classification of Wolbachia strains using wsp gene se-
bachia in host-parasitoid associations. Mol. Biol. Evol. 16: 1711–
quences. Proc. R. Soc. Lond. B Biol. Sci. 265: 509–515.
Vavre, F., P. Fouillet and F. Fleury, 2003
HIGHLIGHTS OF PRESCRIBING INFORMATION ter with caution to patients with active digestive system disease. (5.8) • Immunizations: Avoid live vaccines. (5.9) These highlights do not include all the information needed to use mycophenolic acid delayed-release tablets safely and effectively. See full prescribing information for • Patients with Hereditary Deficiency of Hypoxanthine-guanine P
PRESENTACIÓN Ponemos a disposición de los lectores la edición de la ALERTA PERIODISTICA correspondiente a agosto de 2010. Como es sabido, se trata de una recopilación de las principales informaciones de carácter económico divulgadas por la prensa nacional durante el mes anterior. En este caso, un rápido vistazo al contenido nos dice que los principales tópicos que constituyeron not
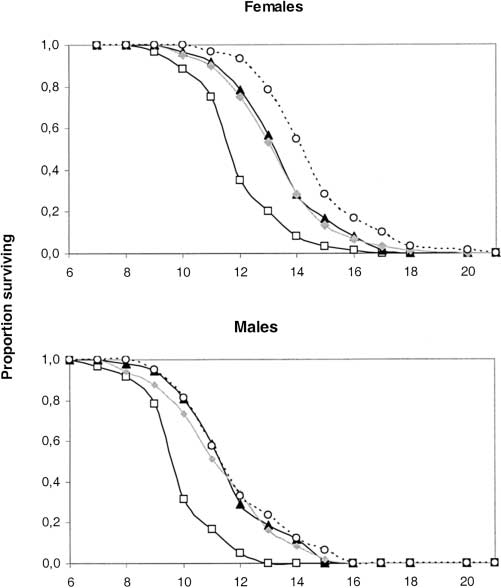
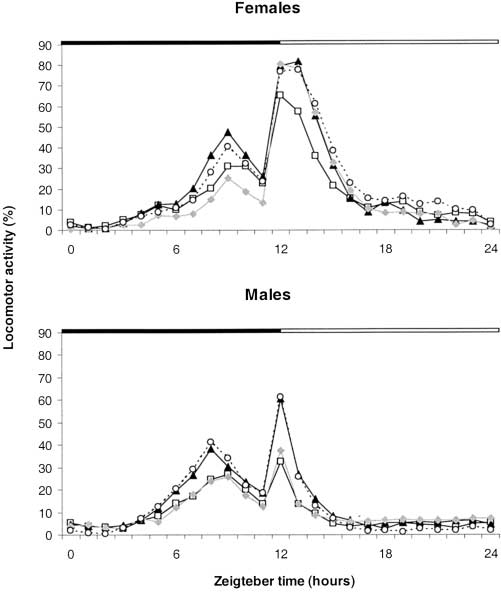 Figure 1.—Starvation survival curves of female and male
Figure 2.—Mean curves of the locomotor activity rhythms
wasps with differing Wolbachia infection statuses. (ᮀ) Pi(123),
of A. tabida lines Pi(123), Pi(13), Pi(23), and Pi(3). Males and
(᭡) Pi(13), (᭜) Pi(23), (᭺) Pi(3).
Figure 1.—Starvation survival curves of female and male
Figure 2.—Mean curves of the locomotor activity rhythms
wasps with differing Wolbachia infection statuses. (ᮀ) Pi(123),
of A. tabida lines Pi(123), Pi(13), Pi(23), and Pi(3). Males and
(᭡) Pi(13), (᭜) Pi(23), (᭺) Pi(3).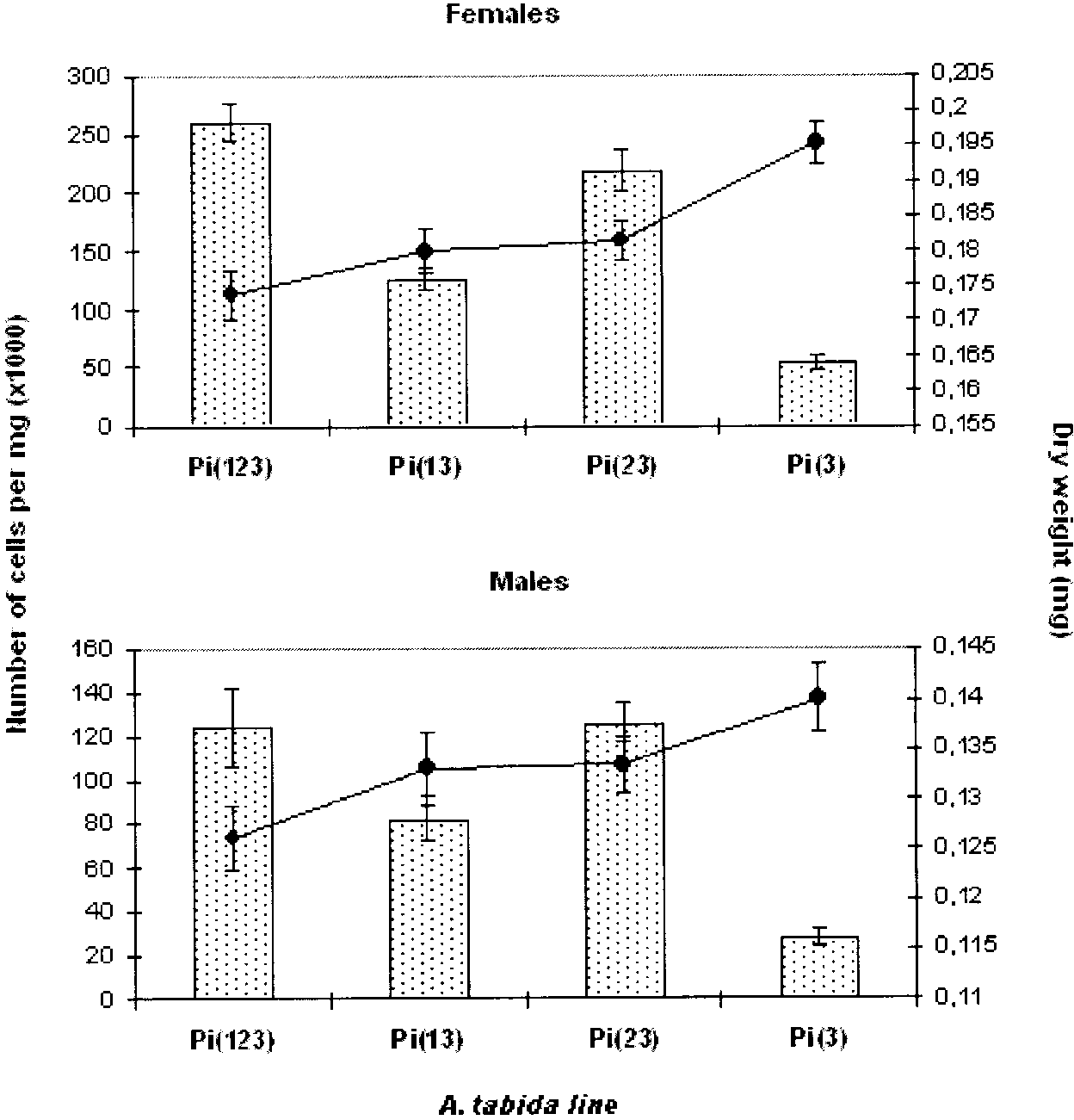 Virulence, Multiple Infections and Regulation of Symbiotic Population
Wolbachia density in A. tabida species
Virulence, Multiple Infections and Regulation of Symbiotic Population
Wolbachia density in A. tabida species Figure 4.—Specific densities of wAtab1, wAtab2, and wAtab3. Specific densities of wAtab1, wAtab2, and wAtab3 in females and
males of various infection statuses are presented. Values correspond to the average of eight individuals per sex and line. Barsshow the standard error. P-values of Kruskal-Wallis tests are indicated. For wAtab3, means marked with the same letters are notsignificantly different (Mann-Whitney tests, P ϭ 0.05).
Figure 4.—Specific densities of wAtab1, wAtab2, and wAtab3. Specific densities of wAtab1, wAtab2, and wAtab3 in females and
males of various infection statuses are presented. Values correspond to the average of eight individuals per sex and line. Barsshow the standard error. P-values of Kruskal-Wallis tests are indicated. For wAtab3, means marked with the same letters are notsignificantly different (Mann-Whitney tests, P ϭ 0.05).