Le métronidazole (Flagyl) reste la référence dans le traitement des infections anaérobies et des parasitoses comme la giardiase ou l’amibiase. Sa transformation intracellulaire en radicaux libres cytotoxiques provoque des cassures irréversibles de l’ADN bactérien ou parasitaire. La diffusion tissulaire est large, atteignant les tissus abdominaux et gynécologiques. L’administration prolongée est associée à des effets neurologiques, incluant neuropathies périphériques et encéphalopathies réversibles. L’association avec l’alcool déclenche une réaction de type antabuse. Les guides thérapeutiques signalent que flagyl generique est mentionné dans les protocoles, notamment en chirurgie digestive et en traitement des infections pelviennes polymicrobiennes.
Immundefekt.eu
CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY, Jan. 2001, p. 74–78
1071-412X/01/$04.00ϩ0 DOI: 10.1128/CDLI.8.1.74–78.2001
Copyright 2001, American Society for Microbiology. All Rights Reserved.
Apoptosis in T-Lymphocyte Subsets in Human Immunodeficiency
Virus-Infected Children Measured Immediately Ex Vivo
TIM NIEHUES,1,2 THOMAS W. MCCLOSKEY,1 JENNIFER NDAGIJIMANA,2 GERD HORNEFF,2
Department of Pediatrics, Division of Allergy and Immunology, North Shore University Hospital/New YorkUniversity School of Medicine, Manhasset, New York, and Children’s Hospital,Heinrich Heine University, Du¨sseldorf, Germany2
Received 2 May 2000/Returned for modification 21 August 2000/Accepted 5 October 2000
Phosphatidylserine molecules are translocated to the outer plasma membrane of lymphocytes undergoing apoptosis and can be detected by the binding of fluorochrome-conjugated annexin V. Using the annexin V assay, we examined CD4 and CD8 T cells from human immunodeficiency virus (HIV)-infected children for apoptosis upon isolation or following in vitro culture. Immediate ex vivo analysis or overnight culture showed significantly higher levels of apoptosis in CD8 cells than in CD4 cells. Following culture with the activating stimulus phytohemagglutinin or anti-CD3 monoclonal antibody, we observed an increase in the percentage of apoptotic CD4 cells, whereas there was no change in the rate of CD8 cell death. These results demonstrate that in HIV-infected children, CD8 apoptosis may occur at a greater rate than CD4 apoptosis in vivo; greater CD4 depletion may be observed due to more efficient mechanisms for peripheral lymphocyte replacement in the CD8 compartment. Furthermore, our data suggest that CD8 lymphocytes may be maximally activated in vivo, a condition which may lead to the exhaustion of CD8-mediated immunity. These findings clarify the differences between the CD4 and CD8 apoptotic responses to HIV.
While an increased rate of lymphocyte apoptosis has been
idate the specificity of annexin binding for apoptosis, we
documented for patients with human immunodeficiency virus
examined whether PBMC which bound annexin V simulta-
(HIV) infection (20), the precise mechanism(s) is still unclear.
neously demonstrated DNA strand breaks by the terminal
Different pathways leading to apoptosis have been proposed;
deoxynucleotidyltransferase-mediated dUTP nick end label-
there is evidence for direct cytopathic effects by viral compo-
ing (TUNEL) method. We found that the percentage of CD8 T
nents as well as indirect effects on bystander cells (9, 11, 18,
lymphocytes undergoing apoptosis was greater than that of
19). While both CD4 and CD8 T cells undergo apoptosis, the
CD4 cells when measured immediately ex vivo or following
induction and kinetics of cell death may be different for each
overnight culture. Furthermore, the addition of activating
subset. For example, telomeres have been observed to be sig-
stimuli in vitro was able to increase the percentage of CD4 cells
nificantly shorter in CD8 cells than in CD4 cells of HIV-
which were dying, whereas the CD8 subset was unchanged.
infected adults, suggesting faster turnover in the CD8 popula-
tion (7, 21, 24). In addition, in vitro addition of interleukin-2
MATERIALS AND METHODS
failed to rescue activated CD8 lymphocytes undergoing apo-
ptosis (15), indicating that these cells may be committed to
Study subjects. Peripheral blood samples were obtained from 67 children with
death in vivo. The deletion of activated responding CD8 T
perinatal HIV infection. Children were grouped using Centers for Disease Con-trol and Prevention classification by the level of immune suppression: category 1,
lymphocytes following an infection may be a homeostatic pro-
none (n ϭ 23, age ϭ 8.2 Ϯ 3.7 years [mean Ϯ standard deviation]); category 2,
cess serving to restore normal cell numbers, an event which
moderate (n ϭ 24, age ϭ 7.5 Ϯ 4.0 years); category 3, severe (n ϭ 20, age ϭ
may be amplified due to the chronic nature of HIV infection.
11.3 Ϯ 5.6 years). Treatments consisted of no antiretroviral therapy (n ϭ 6),
Together, these data suggest that during HIV infection, CD8
reverse transcriptase inhibitor therapy (n ϭ 40), or combination therapy withreverse transcriptase inhibitors and protease inhibitors (n ϭ 21). In a subset of
cells primarily undergo activation-induced cell death due to an
patients for whom viral load measurements were available (n ϭ 46), the median
environment of persistent inflammation.
number of RNA copies per ml was 2,050 (25th to 75th percentile, 400 to 11,000).
Cells undergoing apoptosis translocate phosphatidylserine
Control blood samples were obtained from 10 HIV-negative healthy children
(PS) to their outer cell membrane (23). Annexin V, which
(age ϭ 3.5 Ϯ 2.9 years). These children were free of infection and were under-
binds PS, was used to investigate lymphocyte subset apoptosis
going elective surgery for nonmalignant disorders (inguinal hernia or phimosis). In all cases, informed consent was obtained from the parents or guardians of the
in a cohort of HIV-positive children and uninfected, healthy
children per institutional review board-approved protocols.
pediatric controls. Importantly, the method of annexin V la-
Cell isolation and culture. Following collection into heparinized tubes, sepa-
beling allowed us to quantitate apoptosis in peripheral blood
ration of mononuclear cells was performed by conventional Ficoll-Hypaque
mononuclear cells (PBMC) directly after isolation. To val-
(Lymphoprep; Nycomed AS, Oslo, Norway) density gradient centrifugation. Identical conditions were used for samples from HIV-infected children andhealthy uninfected controls. Each sample was processed within 1 h of collection. In some experiments, PBMC were cultured overnight at a concentration of
* Corresponding author. Mailing address: North Shore University
106/ml at 37°C and 5% CO2 in RPMI 1640 (Gibco Laboratories, Grand Island,
Hospital, 350 Community Drive, Manhasset, NY 11030. Phone: (516)
N.Y.) supplemented with 10% heat-inactivated fetal calf serum (Gibco) and
562-4641. Fax: (516) 562-2866. E-mail: spahwa@nshs.edu.
2 mmol of L-glutamine (Whittaker Bioproducts, Walkersville, Md.) per liter with
EX VIVO APOPTOSIS DURING PEDIATRIC HIV INFECTION
FIG. 1. Flow cytometric gating strategy for determination of total lymphocyte apoptosis and demonstration of simultaneous application of
annexin V and TUNEL assays. Apoptotic lymphocytes (quadrant D2, 9%) which exhibited DNA strand breaks (TUNEL assay) also expressed PS
on their cell membrane (annexin V assay). Monocytes bound annexin V despite the absence of strand breaks (quadrant E1). To accurately assess
total lymphocyte apoptosis (quadrant F2, 17%), a two-tiered strategy of a large light scatter gate (gate B) combined with exclusion of CD14ϩ cells
100 U of penicillin G per ml and 100 g of streptomycin per ml. For determi-
those obtained with the large gate (22, 26, and 55%, respec-
nation of the effect of activation on lymphocyte apoptosis, PBMC (106/ml) were
tively). Worthy of note is our observation that all CD14ϩ cells
incubated overnight with phytohemagglutinin (PHA; 1 g/ml) or anti-CD3monoclonal antibody (0.1 mg/ml, clone HIT 3a; Pharmingen, San Diego, Calif.).
(monocytes) bound annexin V but most were TUNEL nega-
Verification of annexin V assay. PBMC were cultured overnight and then
tive, a finding whose implications are at present unknown. One
labeled with phycoerythrin (PE)-conjugated anti-CD14 monoclonal antibody
possible explanation is that monocytes bind membrane frag-
(Becton Dickinson, San Jose, Calif.), biotinylated annexin V (Pharmingen), and
ments of apoptotic cells which contain PS. Such a situation
streptavidin allophycocyanin (Molecular Probes, Eugene, Oreg.). Samples werefixed with Permeafix reagent (Ortho, Raritan, N.J.) for 40 min at room temper-
would enable the monocytes to become labeled with annexin.
ature, followed by incubation with TUNEL solution as directed by the manufac-
In contrast, the apoptotic lymphocytes which expressed PS as
turer (Phoenix Flow Sytems, San Diego, Calif.). Positive- and negative-control
determined by annexin V binding also exhibited DNA strand
samples were prepared for each experiment. Samples were stored at 4°C until
breakage by the TUNEL assay (Fig. 1). These preliminary
flow cytometric analysis was performed.
findings allowed optimization of our gating for accurate mea-
Quantification of apoptosis within defined lymphocyte populations. For de-
termination of apoptosis within specific cell populations, samples were labeled
surement of lymphocyte apoptosis in subsequent experiments.
with PE-conjugated monoclonal antibodies directed against CD4 or CD8 (Bec-
Apoptosis measured immediately ex vivo in T-cell subsets.
ton Dickinson). Background fluorescence was determined with isotype-matched
Since our objective was to account for all apoptotic lympho-
control antibodies. Immediately after isolation or following overnight culture,
cytes within a sample, the large gate was used with monocytes
PBMC were incubated at room temperature for 10 min with monoclonal anti-bodies and with annexin V conjugated to fluorescein (Boehringer Mannheim,
excluded by gating on CD4bright or CD8bright cells, which limits
Mannheim, Germany). Annexin binding buffer (Pharmingen) was used for all
analysis to T lymphocytes. In order to determine the in vivo
washes and incubation steps. Samples were analyzed by flow cytometry immedi-
levels of cell death, we measured apoptosis immediately after
ately after being washed. The time interval between blood draw and flow cyto-
phlebotomy (Fig. 2) in CD4 and CD8 cells isolated from chil-
metric analysis was less than 2 h in all experiments. Statistical analysis. Differences between groups were determined in a paired
dren of immune categories 1, 2, and 3 as well as from healthy
manner using the paired Student t test or the Wilcoxon signed rank test as
uninfected control children. In HIV-positive children from all
appropriate, depending on the normality of the data distribution, with Excel 5.0
immune categories, apoptosis was significantly higher in CD8
software (Microsoft, Redmond, Wash.) or Sigmastat software (Jandel Scientific,
cells than in CD4 cells (Fig. 3). The percentage of apoptotic
San Rafael, Calif.). P values below 0.05 were considered statistically significant.
CD4 cells was significantly higher than that of controls only inchildren with severe disease, while the percentage of apoptotic
CD8 cells was significantly increased in all infected children. Validation of annexin V assay. Our initial experiments were
As the majority of the patients were receiving antiretroviral
designed to verify results obtained with the annexin assay in
therapy at the time of this cross-sectional study, it was not
our system. Additionally, we investigated appropriate flow cy-
possible to evaluate the effects of treatment on apoptosis.
tometric gating schemes, taking into account the effect of in-
However, elevated levels of CD8 T-lymphocyte death existed
clusion of monocytes on the measurement of total lymphocyte
despite effective control of viremia.
apoptosis. Cells that undergo apoptosis acquire morphological
Apoptosis in T-cell subsets after in vitro activation. To fur-
alterations which are evident upon flow cytometric analysis as
ther investigate the observed differences in cell death levels
changes in light scatter patterns. As lymphocytes become apo-
between CD4 and CD8 lymphocytes, apoptosis was examined
ptotic, they no longer fall within a typical lymphocyte cluster
in T-cell subsets of HIV-infected children after overnight cul-
but can be differentiated from monocytes by lack of expression
ture in the absence or presence of activation stimuli (Fig. 4).
of CD14 antigen (Fig. 1). We found that a large light scatter
Upon overnight incubation with PHA, CD4 cells from HIV-
gate contained significantly more apoptotic lymphocytes than a
infected children showed an increase in the percentage of
small gate, as values obtained for the percentages of apoptosis
apoptotic cells, whereas the CD8 population did not change
using the small gate for cells from children of categories 1, 2
(Fig. 5). In cells from uninfected children, the percentage of
and 3 (9, 10, and 14%, respectively) were always lower than
apoptotic CD4 and CD8 cells did not change after overnight
FIG. 2. Representative histograms demonstrating apoptosis measured immediately ex vivo in T-cell subsets. Samples from healthy children
(CONT) or HIV-infected children (HIVϩ) were labeled with anti-CD4 (A) or anti-CD8 (B) PE and annexin V fluorescein isothiocyanate
immediately upon isolation. Flow cytometric analysis consisted of a combination of a large light scatter gate and a gate on CD4bright or CD8bright
cells (not shown). The percentages of apoptotic CD4 T cells (control, 8%; HIV positive, 18%) and apoptotic CD8 T cells (control, 11%; HIV
positive, 53%) were then determined.
stimulation with PHA. The addition of anti-CD3 monoclonal
antibody also induced an increased percentage of apoptosis in
CD4 T cells from HIV-infected children, whereas apoptosis in
CD8 lymphocytes did not change (data not shown). DISCUSSION
While the ability of HIV to induce lymphocyte cell death is
well established, the causative mechanism(s) remains elusive.
Putative pathways include induction via Fas ligand (2) or tu-
mor necrosis factor-related apoptosis-inducing ligand (14) and
loss of protective molecules such as Bcl-2 (3). Results of the
analysis of T-cell apoptosis in freshly isolated blood samples
from HIV-infected persons most likely reflect the rate of cell
death in vivo, thus providing valuable insight into the patho-
genesis of this disease. Previous reports of immediate ex vivo
apoptosis in HIV-infected adults (12, 13, 16) and children (6),
including a recent study which utilized the annexin V assay
(16), indicated low levels of cell death. In contrast, a study
utilizing the dye Apostain, which is reported to detect early
apoptotic cells, found that over half of freshly isolated lympho-
FIG. 3. Summary of immediate ex vivo apoptosis percentages in
cytes underwent apoptosis in HIV-infected adults (1). These
T-cell subsets. Mean percentages Ϯ standard deviations of annexin
differences may be explained by the patient cohorts studied but
binding CD4 and CD8 cells from HIV-infected children in immune
are more likely due to the apoptosis assays used and the anal-
categories 1 (n ϭ 10), 2 (n ϭ 10), and 3 (n ϭ 11) and from healthy
ysis schemes employed. Our experiments were designed to
controls (n ϭ 10) directly after isolation are shown. Asterisks indicate
statistically significant differences between the percentages of apopto-
account for all apoptotic lymphocytes while omitting poten-
tic CD4 and CD8 cells within each immune category: ء, P Ͻ 0.05; ءء,
tially confounding monocytes from the analysis, and thus our
P Ͻ 0.01; ءءء, P Ͻ 0.001. Differences between groups were determined
results demonstrate higher levels of cell death than many pre-
in a paired manner using the paired Student t test or the Wilcoxon
vious reports. Indeed, levels of cell death observed in a con-
signed rank test. In addition, both CD4 and CD8 apoptosis levels in
ventional lymphocyte gate were of a magnitude similar to those
patients from category 3 were significantly higher than those from
category 1 or 2 (for CD4, P was 0.004 for categories 1 and 3 and 0.01
previously reported, indicating that a basic variable such as
for categories 2 and 3; for CD8, P was 0.004 for categories 1 and 3 and
gating can have tremendous influence on the level of apoptosis
EX VIVO APOPTOSIS DURING PEDIATRIC HIV INFECTION
FIG. 4. Representative histograms demonstrating apoptosis in T-cell subsets after activation. Samples from a healthy child (A) and an
HIV-infected child (B) were cultured overnight in the presence or absence of PHA. Cells were then labeled with anti-CD4 or anti-CD8 PE and
annexin V fluorescein isothiocyanate. Flow cytometric analysis consisted of gating on CD4bright or CD8bright events and quantifying the percentages
of CD4 control apoptotic cells (resting, 8%; activated, 10%), CD8 control apoptotic cells (resting, 21%; activated, 21%), CD4 HIV-positive
apoptotic cells (resting, 18%; activated, 38%), and CD8 HIV-positive apoptotic cells, (resting, 46%; activated, 59%).
observed. We observed that in the majority of samples from
HIV-infected children, CD8 apoptosis was significantly higher
than in controls at all disease stages and apoptosis was present
at higher levels in the CD8 population than in the CD4 pop-
ulation. Because an earlier report of lymphocyte apoptosis in
children (6) indicated that levels of immediate ex vivo CD4 and
CD8 cell death were similar, we suggest that our effort to
include all apoptotic lymphocytes in our analysis may explain
The percentages of peripheral blood CD4 apoptosis were
elevated only in children in immune category 3. Since most
children were on antiretroviral therapies with moderate to
complete virus suppression, relatively low CD4 apoptosis levels
may reflect the adequacy of virus control. Alternatively, CD4
lymphocyte death may predominantly occur in locations other
than the peripheral circulation. However, in agreement with
our findings, in experiments conducted with tonsillar tissues
from HIV-infected adults, Rosok and coworkers found higher
levels of cell death in the CD8 population than in the CD4
population (22). CD8 lymphocyte death during HIV infection
has been associated with cellular activation resulting in in-
creased sensitivity to apoptosis (3, 5). The physiologic process
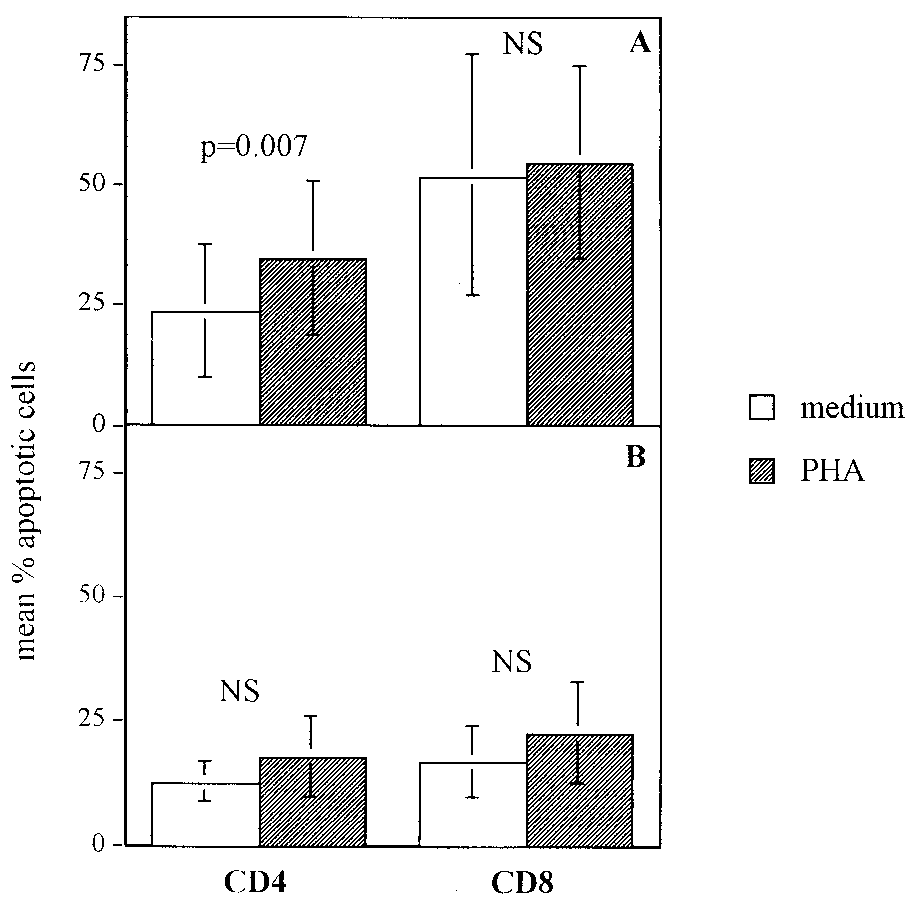
FIG. 5. Summary of percentages of apoptosis in T-cell subsets fol-
lowing activation. Shown are mean percentages Ϯ standard deviations
of response to a pathogen, i.e., activation, may lead to a greater
of annexin binding CD4 and CD8 cells of HIV-infected children (n ϭ
propensity for CD8 death, a pathway which may be augmented
31) (A) and uninfected controls (n ϭ 10) (B) after overnight culture
by the chronic nature of HIV. CD8 T lymphocytes from pa-
with PHA or medium. For HIV-infected CD4 cells, the difference
tients with acute Epstein-Barr and varicella-zoster virus infec-
between the percentage of apoptosis with overnight culture in medium
tions have been demonstrated to be highly sensitive to apopto-
and that with overnight culture in PHA was statistically significant (P ϭ
0.007). Differences between groups were determined in a paired man-
sis (4). Elevated levels of CD8 cell death may thus be a
ner using the paired Student t test or the Wilcoxon signed rank test.
common feature of the immune system’s response to viral
challenge. However, evidence which suggests that the CD8
telomeres in the expanded CD28-CD8ϩ cell subset in HIV disease implicate
compartment may inherently possess a greater regenerative
replicative senescence in HIV pathogenesis. AIDS 10:F17–F22.
8. Estaquier, J., T. Idziorek, F. DeBels, F. Barre-Sinousi, B. Hurtrel, A. M.
capacity exists. In studies of lymphocyte reconstitution follow-
Aubertin, A. Venet, M. Mehtali, E. Muchmore, P. Michel, Y. Mouton, M.
ing cancer chemotherapy, Mackall and colleagues showed that
Girard, and J. C. Ameisen. 1994. Programmed cell death and AIDS: signif-
the CD8 pool had returned to baseline by 3 months post-
icance of T cell apoptosis in pathogenic and nonpathogenic primate lentiviral
infections. Proc. Natl. Acad. Sci. USA 91:9431–9435.
therapy, when the CD4 population was only at one-third of its
9. Finkel, T. H., G. Tudor-Williams, N. K. Banda, M. F. Cotton, T. Curiel, C.
starting value (17). Furthermore, in HIV-infected individuals,
Monks, T. W. Baba, R. M. Ruprecht, and A. Kupfer. 1995. Apoptosis occurs
the fraction of proliferating CD8, but not CD4, T lymphocytes
predominantly in bystander cells and not in productively infected cells of
HIV- and SIV-infected lymph nodes. Nat. Med. 1:129–134.
has been reported to be increased (10). Thus, massive amounts
10. Fleury, S., R. J. de Boer, G. P. Rizzardi, K. C. Wolthers, S. A. Otto, C. C.
of CD8 turnover may occur during HIV infection, but the
Welbon, C. Graziosi, C. Knabenhans, H. Soudeyns, P. A. Bart, S. Gallant, J. M. Corpataux, M. Gillet, P. Meylan, P. Schnyder, J. Y. Meuwly, W.
relative levels appear unperturbed, possibly due to their pro-
Spreen, M. P. Glauser, F. Miedema, and G. Pantaleo. 1998. Limited CD4 T
duction and/or proliferation. In an important study differenti-
cell renewal in early HIV infection: effect of highly active antiretroviral
ating the disease-causing abilities of immunodeficiency viruses,
therapy. Nat. Med. 7:794–801.
11. Herbein, G., C. Van Lint, J. L. Lovett, and E. Verdin. 1997. Distinct mech-
in which pathogenic and nonpathogenic strains of simian im-
anisms trigger apoptosis in human immunodeficiency virus type I-infected
munodeficiency virus were compared, CD8 apoptosis was de-
and in uninfected bystander T lymphocytes. J. Virol. 72:660–670.
tected in all cases, whereas CD4 cell death was limited to
12. Johnson, N., and J. M. Parkin. 1998. Antiretroviral therapy reverses HIV
associated abnormalities in lymphocyte apoptosis. Clin. Exp. Immunol. 113:
pathogenic infections (8). Limiting levels of virus via effective
therapy may serve to decrease induction of apoptosis in the
13. Karmochkine, M., C. Parizot, V. Calvez, A. Coutellier, S. Herson, P. Debre,
CD4 pool. These findings, together with our results, put forth
and G. Gorochov. 1998. Susceptibility of PBMC to apoptosis is correlated to
plasma HIV load. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 17:
the notion that one difference between HIV and other chronic
viral infections may be its unique ability to induce death in the
14. Katsikis, P. D., M. E. Garcia-Ojeda, J. F. Torres-Roca, I. M. Tijoe, C. A. Smith, L. A. Herzenberg, and L. A. Herzenberg. 1997. Interleukin-1 con-
verting enzyme-like protease involvement in Fas-induced and activation-
induced peripheral blood T cell apoptosis in HIV infection: TNF-related
apoptosis-inducing ligand can mediate activation-induced T cell death in
ACKNOWLEDGMENTS
HIV infection. J. Exp. Med. 186:1365–1372.
We thank Maria Marecki, Regina Kowalski, Anette Seibt, and Nico
15. Lewis, D. E., D. S. N. Tang, A. Adu-Oppong, W. Schober, and J. R. Rodgers.
1995. Anergy and apoptosis in CD8 T cells from HIV infected persons,
Vente for excellent technical assistance, Suren Chavan and Claus
J. Immunol. 153:412–420.
Meier for many helpful discussions, and Saroj Bakshi for assistance in
16. Lewis, D. E., D. S. N. Tang, X. Wang, and C. Kozinetz. 1999. Costimulatory
pathways mediate monocyte dependent lymphocyte apoptosis in HIV. Clin.
This work was supported by NIH grants AI28281, HD37345, and
Immunol. 90:302–312.
DA05161 and by a research grant from MSD Merck, “HIV Stipendium
17. Mackall, C. L., T. A. Fleisher, M. R. Brown, M. P. Andrich, C. C. Chen, I. M. Feuerstein, I. T. Magrath, L. H. Wexler, D. S. Dimitrov, R. E. and Gress.
1997. Distinctions between CD8 and CD4 T cell regenerative pathways result
in prolonged T cell subset imbalance after intensive chemotherapy. Blood
REFERENCES 89:3700–3707.
1. Aries, S. P., K. Weyrich, B. Schaaf, F. Hansen, R. H. Dennin, and K. Dalhoff.
18. McCloskey, T. W., M. Ott, E. Tribble, S. A. Khan, S. Teichberg, M. O. Paul,
1998. Early T cell apoptosis and Fas expression during antiretroviral therapy
S. Pahwa, E. Verdin, and N. Chirmule. 1997. Dual role of HIV Tat in
in individuals infected with HIV. Scand. J. Immunol. 48:86–91.
regulation of apoptosis in T cells. J. Immunol. 158:1014–1019.
2. Badley, A. D., J. A. McElhinny, P. J. Leibson, D. H. Lynch, M. R. Alderson,
19. Newell, M. K., L. J. Haughn, C. R. Maroun, and M. H. Julius. 1990. Death and C. V. Paya. 1996. Upregulation of Fas ligand expression by human
of mature T cells by separate ligation of CD4 and the T-cell receptor for
immunodeficiency virus in human macrophages mediates apoptosis of unin-
antigen. Nature 347:286–288.
fected T lymphocytes. J. Virol. 70:199–206.
20. Oyaizu, N., T. W. McCloskey, M. Coronesi, N. Chirmule, V. S. Kalyanara-
3. Bofill, M., W. Gombert, N. J. Borthwick, A. N. Akbar, J. E. McLaughlin, man, and S. Pahwa. 1993. Accelerated apoptosis in peripheral blood mono- C. A. Lee, M. A. Johnson, A. J. Pinching, and G. Janossy. 1995. Presence of
nuclear cells (PBMCs) from human immunodeficiency virus type-1 infected
CD3ϩCD8ϩBcl-2low lymphocytes undergoing apoptosis and activated mac-
patients and in CD4 cross-linked PBMCs from normal individuals. Blood
rophages in lymph nodes of HIV patients. Am. J. Pathol. 146:1542–1555. 82:3392–3400.
4. Borthwick, N. J., M. Bofill, I. Hassan, P. Panayiotidis, G. Janossy, M.
21. Palmer, L. D., N. Weng, B. L. Levine, C. H. June, H. C. Lane, and R. J. Salmon, and A. N. Akbar. 1996. Factors that influence activated CD8 T cell Hodes. 1997. Telomere length, telomerase activity, and replicative potential
apoptosis in patients with acute herpesvirus infections: loss of costimulatory
in HIV infection: analysis of CD4ϩ and CD8ϩ T cells from HIV-discordant
molecules CD28, CD5, and CD6 but maintenance of bax and bcl-x expres-
monozygotic twins. J. Exp. Med. 185:1381–1386.
sion. Immunology 88:508–515.
22. Rosok, B. I., J. E. Brinchmann, G. Stent, R. Bjerknes, P. Voltersvik, J.
5. Boudet, F., H. Lecoeur, and M. L. Gougeon. 1996. Apoptosis associated with Olofsson, and B. Asjo. 1998. Correlates of apoptosis of CD4 and CD8 T cells
ex vivo down regulation of Bcl-2 and upregulation of Fas in potential cyto-
in tonsillar tissue in HIV infection. AIDS Res. Hum. Retrovir. 14:1635–1643.
toxic CD8 T lymphocytes during HIV infection. J. Immunol. 156:2282–2293.
23. van Engeland, M., L. J. Nieland, C. Ramaekers, B. Schutte, and C. P.
6. Cotton, M. F., D. N. Ikle, E. L. Rapaport, S. Marschner, P. O. Tseng, Reutelingsperger. 1998. Annexin V-affinity assay; a review on an apoptosis R. Kurrle, and T. H. Finkel. 1997. Apoptosis of CD4ϩ and CD8ϩ T cells
detection system based on phosphatidylserine exposure. Cytometry 31:1–9.
isolated immediately ex vivo correlates with disease severity in human im-
24. Wolthers, K. C., G. Bea, A. Wisman, S. A. Otto, A. M. de Roda Husman, N.
munodeficiency virus type 1 infection. Pediatr. Res. 42:656–664. Schaft, F. de Wolf, J. Goudsmit, R. A. Countinho, A. G. van der Zee, L.
7. Effros, R. B., R. Allsopp, C. P. Chiu, M. A. Hausner, K. Hirji, L. Wang, C. B. Meyaard, and F. Miedema. 1996. T cell telomere length in HIV-1 infection: Harley, B. Villeponteau, M. D. West, and J. V. Giorgi. 1996. Shortened
no evidence for increased CD4ϩ T cell turnover Science 274:1543–1547.
EPILEPSY AND YOUR DOG Cheryl L Chrisman DVM, MS, EdS, Diplomate ACVIM-Neurology College of Veterinary Medicine, University of Florida Presented at the University of Florida College of Veterinary Medicine 8th Annual Dog Owners & Breeders Symposium on July 31, 2004. Courtesy of the AKC’s Canine Health Foundation. What is epilepsy? Epilepsy is an electrical disturbance of the
VOLUNTARY STOPPING EATING & DRINKING This paper addresses questions about stopping eating and drinking as an option to hasten death. The U.S. Supreme Court has affirmed the right of a competent individual to refuse medical therapies and this includes food and fluids. This choice is also commonly accepted in the medical community. One of the advantages of this decision is that you may chan
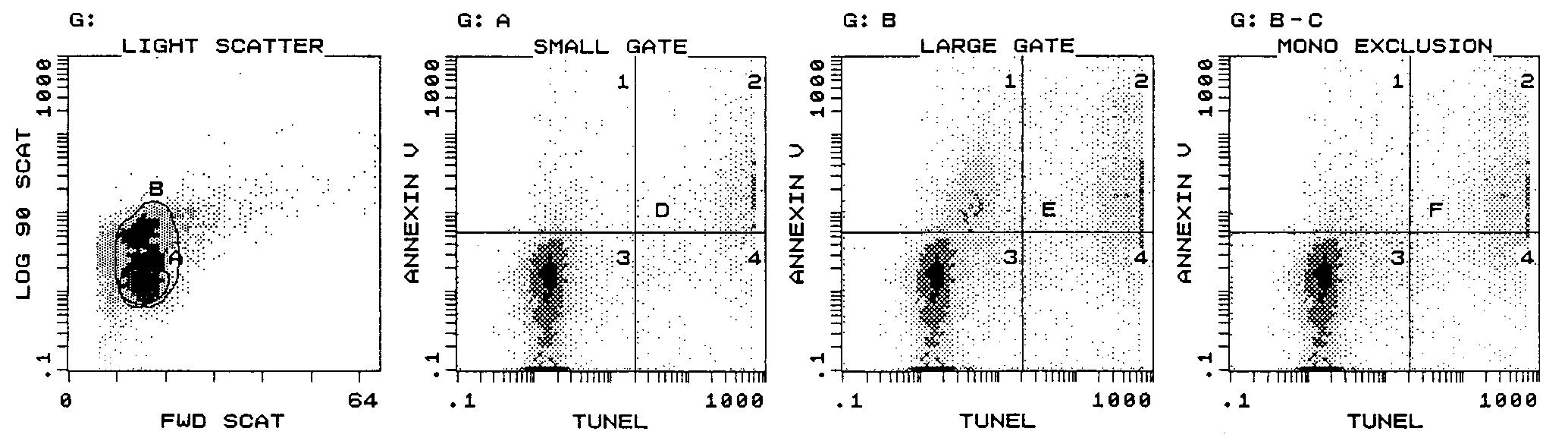 EX VIVO APOPTOSIS DURING PEDIATRIC HIV INFECTION
FIG. 1. Flow cytometric gating strategy for determination of total lymphocyte apoptosis and demonstration of simultaneous application of
annexin V and TUNEL assays. Apoptotic lymphocytes (quadrant D2, 9%) which exhibited DNA strand breaks (TUNEL assay) also expressed PS
on their cell membrane (annexin V assay). Monocytes bound annexin V despite the absence of strand breaks (quadrant E1). To accurately assess
total lymphocyte apoptosis (quadrant F2, 17%), a two-tiered strategy of a large light scatter gate (gate B) combined with exclusion of CD14ϩ cells
100 U of penicillin G per ml and 100 g of streptomycin per ml. For determi-
those obtained with the large gate (22, 26, and 55%, respec-
nation of the effect of activation on lymphocyte apoptosis, PBMC (106/ml) were
tively). Worthy of note is our observation that all CD14ϩ cells
incubated overnight with phytohemagglutinin (PHA; 1 g/ml) or anti-CD3monoclonal antibody (0.1 mg/ml, clone HIT 3a; Pharmingen, San Diego, Calif.).
EX VIVO APOPTOSIS DURING PEDIATRIC HIV INFECTION
FIG. 1. Flow cytometric gating strategy for determination of total lymphocyte apoptosis and demonstration of simultaneous application of
annexin V and TUNEL assays. Apoptotic lymphocytes (quadrant D2, 9%) which exhibited DNA strand breaks (TUNEL assay) also expressed PS
on their cell membrane (annexin V assay). Monocytes bound annexin V despite the absence of strand breaks (quadrant E1). To accurately assess
total lymphocyte apoptosis (quadrant F2, 17%), a two-tiered strategy of a large light scatter gate (gate B) combined with exclusion of CD14ϩ cells
100 U of penicillin G per ml and 100 g of streptomycin per ml. For determi-
those obtained with the large gate (22, 26, and 55%, respec-
nation of the effect of activation on lymphocyte apoptosis, PBMC (106/ml) were
tively). Worthy of note is our observation that all CD14ϩ cells
incubated overnight with phytohemagglutinin (PHA; 1 g/ml) or anti-CD3monoclonal antibody (0.1 mg/ml, clone HIT 3a; Pharmingen, San Diego, Calif.).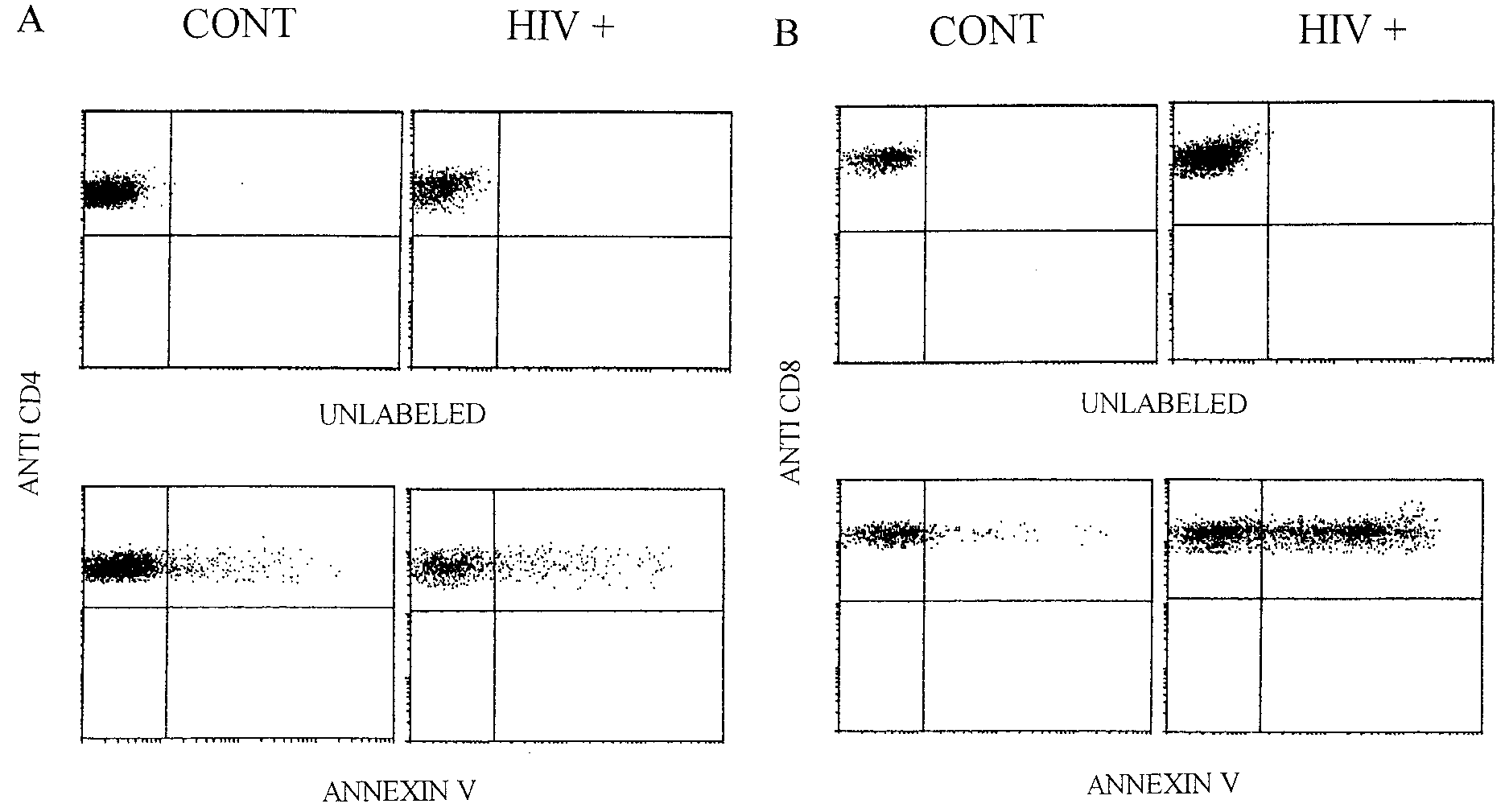
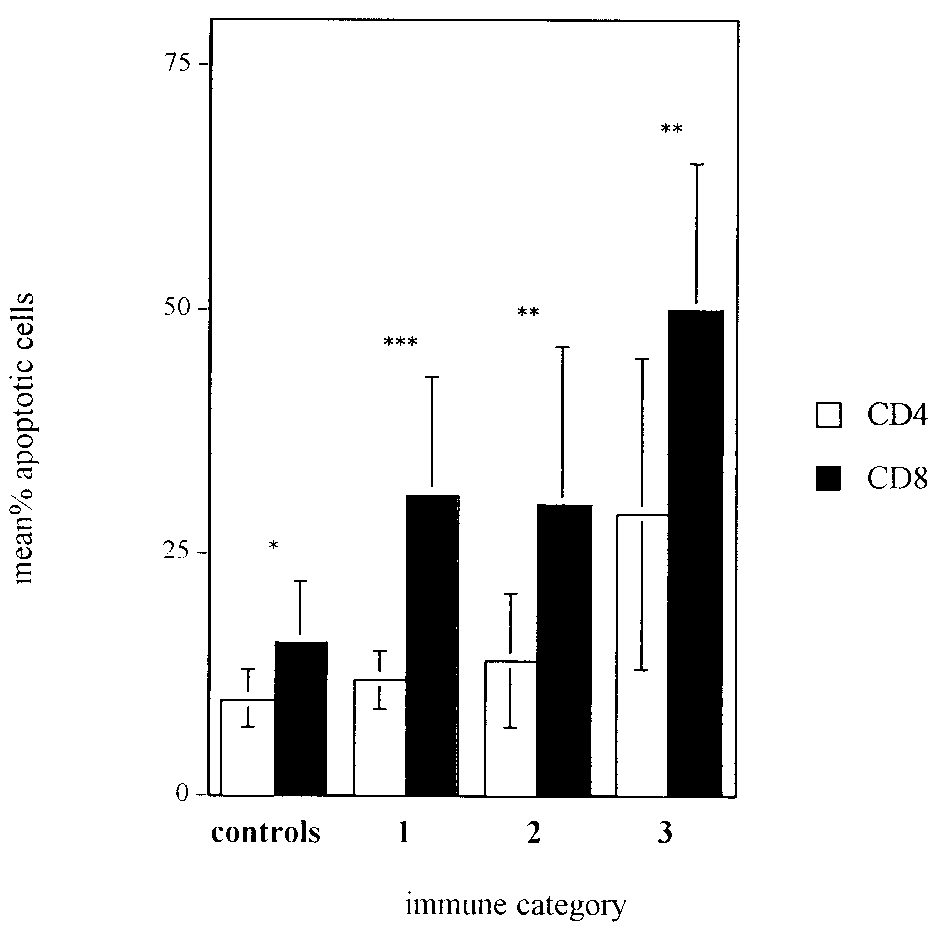 FIG. 2. Representative histograms demonstrating apoptosis measured immediately ex vivo in T-cell subsets. Samples from healthy children
(CONT) or HIV-infected children (HIVϩ) were labeled with anti-CD4 (A) or anti-CD8 (B) PE and annexin V fluorescein isothiocyanate
immediately upon isolation. Flow cytometric analysis consisted of a combination of a large light scatter gate and a gate on CD4bright or CD8bright
cells (not shown). The percentages of apoptotic CD4 T cells (control, 8%; HIV positive, 18%) and apoptotic CD8 T cells (control, 11%; HIV
positive, 53%) were then determined.
FIG. 2. Representative histograms demonstrating apoptosis measured immediately ex vivo in T-cell subsets. Samples from healthy children
(CONT) or HIV-infected children (HIVϩ) were labeled with anti-CD4 (A) or anti-CD8 (B) PE and annexin V fluorescein isothiocyanate
immediately upon isolation. Flow cytometric analysis consisted of a combination of a large light scatter gate and a gate on CD4bright or CD8bright
cells (not shown). The percentages of apoptotic CD4 T cells (control, 8%; HIV positive, 18%) and apoptotic CD8 T cells (control, 11%; HIV
positive, 53%) were then determined.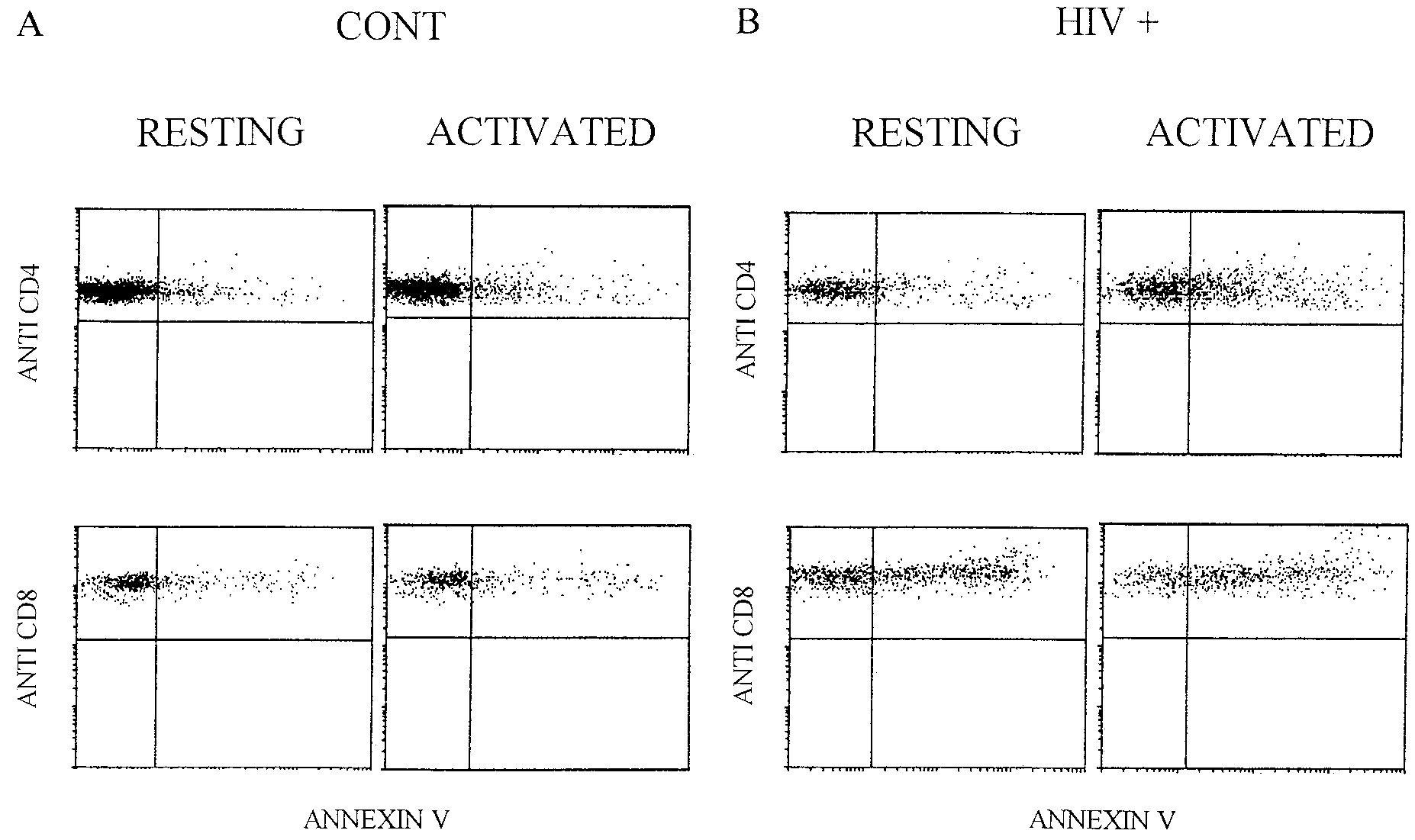
 EX VIVO APOPTOSIS DURING PEDIATRIC HIV INFECTION
FIG. 4. Representative histograms demonstrating apoptosis in T-cell subsets after activation. Samples from a healthy child (A) and an
HIV-infected child (B) were cultured overnight in the presence or absence of PHA. Cells were then labeled with anti-CD4 or anti-CD8 PE and
annexin V fluorescein isothiocyanate. Flow cytometric analysis consisted of gating on CD4bright or CD8bright events and quantifying the percentages
of CD4 control apoptotic cells (resting, 8%; activated, 10%), CD8 control apoptotic cells (resting, 21%; activated, 21%), CD4 HIV-positive
apoptotic cells (resting, 18%; activated, 38%), and CD8 HIV-positive apoptotic cells, (resting, 46%; activated, 59%).
EX VIVO APOPTOSIS DURING PEDIATRIC HIV INFECTION
FIG. 4. Representative histograms demonstrating apoptosis in T-cell subsets after activation. Samples from a healthy child (A) and an
HIV-infected child (B) were cultured overnight in the presence or absence of PHA. Cells were then labeled with anti-CD4 or anti-CD8 PE and
annexin V fluorescein isothiocyanate. Flow cytometric analysis consisted of gating on CD4bright or CD8bright events and quantifying the percentages
of CD4 control apoptotic cells (resting, 8%; activated, 10%), CD8 control apoptotic cells (resting, 21%; activated, 21%), CD4 HIV-positive
apoptotic cells (resting, 18%; activated, 38%), and CD8 HIV-positive apoptotic cells, (resting, 46%; activated, 59%).