Le métronidazole (Flagyl) reste la référence dans le traitement des infections anaérobies et des parasitoses comme la giardiase ou l’amibiase. Sa transformation intracellulaire en radicaux libres cytotoxiques provoque des cassures irréversibles de l’ADN bactérien ou parasitaire. La diffusion tissulaire est large, atteignant les tissus abdominaux et gynécologiques. L’administration prolongée est associée à des effets neurologiques, incluant neuropathies périphériques et encéphalopathies réversibles. L’association avec l’alcool déclenche une réaction de type antabuse. Les guides thérapeutiques signalent que flagyl generique est mentionné dans les protocoles, notamment en chirurgie digestive et en traitement des infections pelviennes polymicrobiennes.
Pq189910428p
Widespread accumulation of [3H]testosterone in the spinal cord of a wild bird with an elaborate courtship display
J. DOUGLAS SCHULTZ AND BARNEY A. SCHLINGER*Department of Physiological Science and Laboratory of Neuroendocrinology, Brain Research Institute, University of California, Los Angeles, CA 90095-1527
Communicated by Peter Marler, University of California, Davis, CA, June 22, 1999 (received for review January 28, 1999)ABSTRACT Elaborate courtship displays are relatively
the performance of these behaviors by birds across many taxa,
common features of the masculine reproductive behavior in
little is known about their hormonal and neural control. Given
birds. However, little is known about their neural and hor-
that some of these displays (i) involve coordinate usage of
monal control. One bird that performs such a display is the
several neuromuscular systems controlling posture and move-
golden-collared manakin (Manacus vitellinus) of Panamanian
ments of the wings, legs, and tail; (ii) are often performed by
forests. Adult males, but not females, perform a physically
males and not females; and (iii) are used in reproductive
intense display requiring substantial neuromuscular control
contexts, we would predict that in displaying birds, the spinal
of the wings and legs. We tested the hypothesis that steroid
motoneurons controlling behaviorally relevant muscles would
sensitivity is a property of neurons in the manakin spinal
be sensitive to steroid hormones and would be anatomically
cord. Males and females were captured from active courtship
and/or physiologically sexually dimorphic. leks, treated with drugs to block steroidogenesis, injected with
To test these hypotheses, we have performed tritiated
3H-labeled testosterone, and the spinal cords were removed
testosterone (3H-T) autoradiography on the spinal cords of
and processed for autoradiography. Sex steroid-accumulating
adult male and female golden-collared manakins (Manacuscells were widely distributed in the spinal cords in each of six vitellinus), a common bird species of central Panamanian
males and in one of five females. Cells, including presumptive
forests. Manakins are a family of suboscine, passerine birds
motoneurons, reached their highest density in the ventral
that are common in forests of the New World tropics. Males
horns of the cervical and lumbosacral enlargements, regions
of several manakin species, including the golden-collared
associated with motor control of the wings and legs. These
manakins, perform elaborate courtship displays involving
results suggest that neurons in the adult manakin spinal cord
short flights with midair acrobatics and intense jumping and
can express sex-steroid receptors, but do so less in females
dancing movements. In addition, the wings of some species
than in males. This evidence for androgen sensitivity and
(including golden-collared manakins) possess sexually dimor-
sexual dimorphism in the adult avian spinal cord suggests that
phic feather structures (14) that assist in producing loud
sex steroids may control diverse behaviors in male birds in
snapping sounds by the rapid flipping of their wings (13, 14). part by acting directly on the spinal neural circuits.
We report that male golden-collared manakins show wide-
spread accumulation of 3H-T or its metabolites in the spinal
To reproduce, males perform behaviors to attract and stimu-
cord, including in many large motoneurons, and this pattern of
late females, defend territories and mates, copulate, and care
sex-steroid accumulation is different in females. Androgen
for young. Sex steroids can control the development and
accumulation in spinal motoneurons in adult birds suggests
expression of many of these behaviors by direct actions on the
that steroid sensitivity may be present in those neural pathways
central nervous system. Many studies focus on the actions of
of the spinal cord that generate a range of avian courtship
sex steroids on the hypothalamus, where there exists a rela-
tively conserved population of neurons expressing androgen
receptors (AR) and estrogen receptors (ER) within circuits
controlling masculine copulatory behaviors (1–3). Steroids can
also act directly on motoneurons (4, 5). For example, AR can
Golden-collared manakins (six males; five females) were cap-
be expressed in mammalian motoneurons of the lumbar spinal
tured in mist nets from active courtship leks located in central
cord that innervate muscles controlling the penis (6–9). By a
Panama in June, 1995 and September, 1996. (All protocols for
combination of actions of estrogens and androgens on the
animal use have been approved by the Chancellor’s Animal
brain and on the spinal cord, male mammals are stimulated to
Research Committee and were collected under permit from
copulate and are functionally able to do so.
Instituto Nacionale de Recursos Naturales Renovables, gov-
Steroids also influence neurons controlling other reproduc-
ernment of Panama.) To reduce endogenous androgen pro-
tive behaviors (4, 10), such as the widely studied neural
duction, birds were injected immediately with an inhibitor of
circuitry controlling song located in the telencephalon of the
one of two steroidogenic enzymes, either trilostane (an inhib-
oscine passerine songbirds (11, 12). Given that song is a
itor of 3--hydroxysteroid dehydrogenase/isomerase, Sterling-
significant acoustic signal coordinating reproduction in these
Winthrop Research Insitute, 2 males and 1 female) or keto-
birds, it is not surprising that the song-control circuitry is
conazole (an inhibitor of 17-␣-hydroxylase/C17–20 lyase, Jans-
influenced by sex steroids via the expression of AR and ER.
sen; 4 males and 4 females). Studies on the effectiveness of
But song is just one of a suite of avian reproductive behaviors.
these two inhibitors in birds are published elsewhere (15, 16).
Male birds can exhibit an impressive repertoire of visual and
After 24 hr (trilostane) or 8–12 hr (ketoconazole), the birds
acoustic reproductive displays (13). In some species, these
were injected with 60–80 Ci (1 Ci ϭ 37 GBq) of 3H-T
visual displays can be dramatic, including acrobatic movements
(specific activity 102.5 Ci/mmol; New England Nuclear) and
that are enhanced by conspicuous physical ornaments. Despite
sacrificed 90 min later by decapitation. The gonads were
The publication costs of this article were defrayed in part by page charge
Abbreviations: AR, androgen receptor; ER, estrogen receptor; 3H-T,
payment. This article must therefore be hereby marked ‘‘advertisement’’ in
*To whom reprint requests should be addressed at: Department of
accordance with 18 U.S.C. §1734 solely to indicate this fact.
Physiological Science, P.O. Box 951527, Los Angeles, CA 90095-
PNAS is available online at www.pnas.org.
1527. E-mail: Schlinge@lifesci.ucla.edu. Proc. Natl. Acad. Sci. USA 96 (1999)
visually inspected and sizes noted. The vertebral columns were
immediately dissected free of surrounding tissues and fixed
briefly (Ϸ1 min) in formalin (3.7% formaldehyde in 0.9%
Sex steroid-accumulating cells were found in the manakin
PBS) to assist with removal of the spinal cord (which remained
spinal cord with substantially more cells in males than in
unfixed). The spinal cords were then cut lengthwise, into halves
females. Accumulation of sex steroid was found in cells of the
spinal cords of all six males (on average, 209 cells over the
or thirds, flash-frozen with crushed dry ice onto cork with
entire cord). By contrast, only one female approached this
Tissue-Tek (Sakura Finetek, Torrance, CA), and stored in dry
number (with 101 cells found). Only four, two, two, and zero
ice at Ϫ80°C. The cords were transported in dry ice to the
accumulating cells per spinal cord were found in the remaining
United States and prepared for autoradiography.
Cords were sectioned longitudinally in the dark on a cryostat
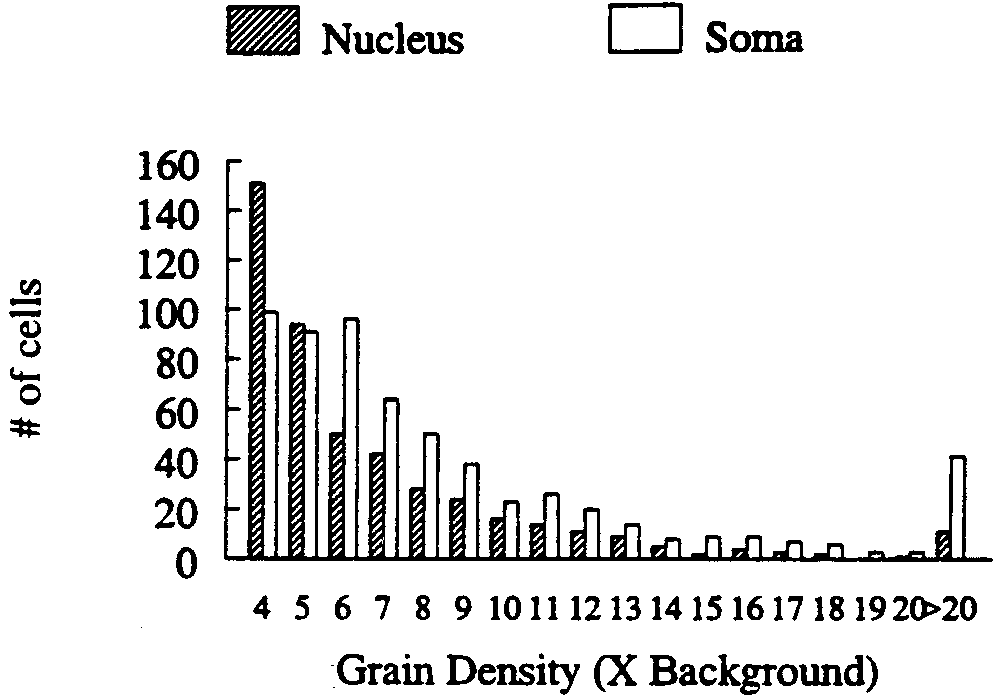
Cells with a visible nucleus comprised 59.5% of the total
(at Ϫ17°C) and thaw-mounted onto microscope slides. For
number of sex steroid-accumulating cells (Fig. 1). Of the
autoradiographic analysis, two consecutive sections were cut at
nucleated cells, 95% had a greater density of silver grains over
6 m and thaw-mounted onto separate slides previously
the nucleus alone as compared with the whole soma (which
dipped in Kodak NTB-2 photographic emulsion (series 1 and
includes the nucleus). Fig. 2 illustrates the total number of
2, respectively). The third consecutive section was sectioned at
nucleated and nonnucleated cells with nuclear or somal accu-
20 m and thaw-mounted on slides (Fisher Superfrost Plus).
mulation (respectively) greater than 3ϫ background. Many
These slides were later stained with thionin and used to assist
cells in males were present with accumulation between 3 and
with preparation of a spinal cord map. The fourth consecutive
5ϫ background (64.0 on average) but were not accepted as
section (cut at 6 m) was usually discarded because of poor
representing significant accumulation based on our criteria. In
quality; this cycle was repeated through the entire cord.
the female in which we found a large number of cells meeting
Although only one series was analyzed from a given bird, there
the 5ϫ criteria, 39 additional cells met the 3ϫ criteria. If these
was usually 38 m of tissue between each section in a given
cells do indeed express AR or ER, but at low levels, then our
series. Slides from series 1 and 2 were sealed in light-tight,
results may underestimate the total number of sex steroid-
desiccated containers and stored at 4°C. After 3, 6, 9, or 12
accumulating cells in the manakin spinal cord (21). No addi-
months, autoradiographic slides were immersed consecutively
tional cells that met the 3ϫ criteria were found in the
in Kodak D-19 developer, distilled H2O, and Kodak fixer,
thionin stained, dehydrated in a graded series of alcohols, and
Sex steroid-accumulating cells with an obvious nucleus fell
into two general size categories, small and large cells with
For each bird, the slide series demonstrating optimal cellu-
mean areas of Ϸ1,000 m2 and 4,000 m2, respectively (Fig. 3).
lar-to-background silver grain density was chosen for analysis.
Most large cells were located ventrally, whereas small cells
Cells that appeared to have high levels of accumulation with
were distributed widely, especially in middle and dorsal levels
a distinct nucleus were analyzed over both the nucleus and
soma (which included the nucleus). Cells that appeared to have
Even though sex steroid-accumulating cells were found in all
high levels of accumulation but lacked an obvious nucleus were
six males, only four males provided histology that was quali-
analyzed for grain accumulation over the whole soma. These
tatively sufficient to allow for precise localization and map-
were measured by capturing the cellular image (at ϫ1,000) on
ping. Sex steroid-accumulating cells of the remaining two
a Macintosh computer by using a Zeiss Axioskop microscope
males were present with their size and position consistent with
linked to a Sony charge-coupled device video camera. By using
the large motoneurons of the ventral cervical and lumbosacral
enlargements. For purposes of description, the spinal cord was
ference of the nucleus, soma, or selected region of neuropil was
subdivided rostrocaudally into the high cervical, cervical en-
traced, and silver grain size was defined. We computed both
largement, midthoracic, and lumbosacral enlargement regions
and dorsoventrally into the ventral, middle, and dorsal regions
the area and the number of silver grains contained within the
(Fig. 4). Cells were included on the map without distinguishing
tracing. Silver-grain density was determined by dividing the
between those with and without nuclei. The relative abun-
area measurement into the number of silver grains counted. A
dance and distribution of sex steroid-accumulating cells in the
similar measurement was then made over the remaining
spinal regions of the four males and five females is summarized
neuropil adjacent to the cell in the captured microscope image
(background). The average background area measured was six
times larger than the average motoneuron somal area. We then
calculated the ratio of silver-grain density over the soma or
nucleus to background density. Data were recorded for all cells
in which this cellular or nuclear-grain density was Ն3 times the
background; accumulation was considered significant when
cellular or nuclear-grain densities were Ն5 times background
(17, 18), and these cells are hereafter referred to as sex
A map of the position of all sex steroid-accumulating cells
in the spinal cord was created for each individual bird;
afterward, a single composite map was created summarizing
the general distribution of all birds. Cellular position in the
rostrocaudal plane was established by observing landmarks
such as visible dorsal-root ganglion, the beginning of the
cervical enlargement that starts at C10 in the pigeon (19), and
the lumbosacral enlargement that starts at L1 in the pigeon
(20). Cells in the dorsoventral axis were located by estimating
the proximity of the cell to Lamina IX (20), a morphologically
FIG. 1. Representative sex steroid-accumulating, thionin-stained
prominent motoneuronal cell column throughout the ventral
neurons from the ventral cervical enlargement of an adult male
Proc. Natl. Acad. Sci. USA 96 (1999)
females in the middle third of the cord, and a group of cells
from one male was found in the dorsal third of the cord.
Sex steroid-accumulating cells were widespread in the lum-
bosacral enlargement (L1–L6) throughout its full rostrocaudal
extent. A majority of these cells were found in the ventral third
of the enlargement and were found in one male and in one
female. Numerous labeled cells were also found in medial
portions of the middle third of the lumbosacral enlargement,
especially in two males. One of these males also had a
significant number of cells in the dorsal third of the enlarge-
ment. We were unable to reliably distinguish the glycogen body
DISCUSSION
These results suggest that some neurons in the manakin spinal
cord express AR as described in other vertebrates (6, 9, 22). It
is also possible that 3H-T was aromatized into [3H]estradiol
FIG. 2. Motoneuronal silver grain density (cellular silver grain
number as a multiple of background silver grain number) over nuclei
and that ER are expressed in the manakin spinal cord, as they
(nucleated cells) and somas (nonnucleated cells) after injection of
are in rats (23). The presence of widespread sex-steroid
3H-labeled testosterone into adult male and female golden-collared
accumulation in cells in the spinal cords of males compared
with females suggests that there is a sex difference in the
magnitude of AR and/or ER expression or in the number of
Within the high cervical cord (rostral to the cervical en-
AR- and/or ER-expressing cells. It is also possible that sex
largement), accumulating cells were found in all four males
steroid availability in the manakin spinal cord may differ
and in none of the females. Here, the number of cells were
between males and females because of sex differences in
roughly evenly distributed dorsoventrally, with the middle
steroid metabolism (24, 25). Presumably, sex steroids act on
third of the high cervical region having the most cells relative
spinal neural circuits in manakins to control expression of
to the ventral and dorsal regions. Localization was mostly
confined to an area midway between the hindbrain and the
Many sex steroid-accumulating cells in the manakin spinal
cervical enlargement (see Fig. 4), approximately C5.
cord are located in lamina IX (20) and are of a large size,
In the cervical enlargement (C10–C13), sex steroid-
consistent with that of motoneurons (Fig. 2). Similar AR-
accumulating cells were abundant in the ventral third for three
expressing motoneurons have been found in rat (23, 26) and in
of the four males and for one female. Relative to the ventral
Xenopus (27), leading us to suspect that these ventral mo-
third, there were comparatively fewer cells in the middle third,
toneurons in manakins are also androgen-, and not estrogen-,
and accumulation was present in all four males and none of the
sensitive. Some smaller sex steroid-accumulating cells found in
females. In the ventral and middle third of the cervical
dorsal regions of the manakin spinal cord may be sensory
enlargement, cells were found along the entire rostrocaudal
neurons binding estrogen as reported in rats (23) and ring
length. In the dorsal third, we found one male with two distinct
doves (28). Subpopulations of AR-expressing motoneurons
clusters of cells, one each in the rostral and caudal portions of
the cervical enlargements. We found no sex steroid-
Table 1. Relative abundance of sex steroid-accumulating cells in
accumulating cells in this region of any other bird.
A small number of sex steroid-accumulating cells were
present in the midthoracic region, approximately T3, midway
between the cervical and lumbosacral enlargements. Only one
male showed a few accumulating cells in the ventral third of the
cord. Small cell clusters were found in two males and two
Areas of nucleated (hatched bars) and nonnucleated (open
bars) sex steroid-accumulating cells in the spinal cords of adult male
M, male; F, female; Ϫ, no cells; ϩ, 1–5 cells; ϩϩ, 16–50 cells; ϩϩϩ,
and female golden-collared manakins.
51–100 cells; ϩϩϩϩ, 101–200 cells; ϩϩϩϩϩ, 201–224 cells. Proc. Natl. Acad. Sci. USA 96 (1999)
steroids act directly on spinal neural circuits to control the
expression of these courtship behaviors. Males, but not fe-
males, court actively, presumably because testosterone circu-
lates at higher levels in males than in females. We suspect that
androgens, or their metabolites, act centrally to increase the
motivation to court and peripherally to increase the neuro-
muscular capacity to perform the displays. Androgens also act
on neurons centrally and peripherally in frogs and rats to
activate masculine reproductive behaviors (5, 30, 31). In the
spinal cords of these species, sex differences in AR expression
in part underlie the observed sex differences in behavior (5).
The basis of the sex differences in sex-steroid binding in the
manakin spinal cord is unknown. The differences could arise
from sex differences in the numbers of some AR-expressing
cells, as observed in the rat (6, 21), in which fewer AR-
expressing motoneurons exist in the female lumbar spinal cord
because females lack two of the target muscles (5). It is unlikely
that the female manakin possesses fewer motoneurons if they
are innervating essential muscles of the wings and legs. It is
more likely that higher levels of circulating testosterone in
male manakins transiently up-regulate spinal AR to a greater
degree than in females, as is presumed to occur in Xenopus
(27). Although we cannot exclude the possibility that some of
the differences we observe reflect permanent sex differences
in sex-steroid binding or in other cellular attributes, we assume
that neuromuscular control of the wings and legs is temporally
adapted for greater use by sex steroids when males are actively
We found considerable variability in the numbers of sex
steroid-accumulating cells across males and the extent to which
androgens were bound by cells (Table 1). Cells were found
most consistently in the cervical enlargement, but even here
one male showed none. This variability in spinal cord steroid
sensitivity may have been produced by differences in circulat-
ing testosterone if males were in different reproductive con-
dition. Some birds were caught in September, a time when
males are beginning to cease courtship activity (14). Our data
indicate that the male with the smallest testes had the lowest
number of sex steroid-accumulating cells, with greater num-
bers of accumulating cells found in males with somewhat larger
testes. Because androgens can directly regulate AR in the
spinal cord (9), males with low plasma testosterone levels
before capture could have displayed less sex-steroid accumu-
FIG. 4. Representative map showing the spatial distribution of sex
lation. No differences in ovary size were observed that account
steroid-accumulating cells in the golden-collard manakin spinal cord.
Each symbol represents three cells, with isolated cells omitted; black
for differences observed across females. Variability might also
dots represent cells found in males; X represents cells found in females
have been produced artificially if trilostane or ketoconazole
(see text for details). The cord is illustrated at three levels; all cells
reduced endogenous androgen synthesis differently across
found within a given level are illustrated: Ventral, the entire cord
birds, creating disproportionate competition for androgen-
ventral to the bifurcation of the ventral horns, including most of lamina
binding sites with endogenous nonradioactive testosterone.
IX and much of lamina VIII of Leonard and Cohen (1975); dorsal, the
We cannot exclude the possibility that different effects of these
entire cord dorsal to the beginning of the bifurcation of the dorsal
drugs on testicular or ovarian steroidogenesis might also have
horns out to their tips, including all of lamina I–IV of Leonard and
contributed to observed differences in spinal cord sex-steroid
Cohen (1975) and part of lamina V; middle, the remaining cord
between the ventral and dorsal levels. Table 1 illustrates individual
differences in the pattern of accumulation.
The identification of steroid-sensitive neural circuits
throughout the spinal cord of the golden-collared manakin
are functionally associated with androgen-dependent muscles
suggests that sex steroids may have a broader role in modu-
and together control androgen-dependent penile reflexes in
lating avian neuromuscular systems than previously thought.
rodents (6, 25, 9) and amplexus in Xenopus (22, 27). Therefore,
Although birds have been widely studied with respect to
we assume that at least some of these sex steroid-accumulating
steroid actions on the brain, no avian neuromuscular system
motoneurons in manakins are also part of androgen-
dependent neuromuscular systems. Because in manakins the
has been fully exploited to evaluate steroid control of mo-
majority of sex steroid-accumulating cells are found in the
toneurons and sexually dimorphic muscles that they might
cervical and lumbosacral enlargements, and motoneurons in
innervate. As in other species, sex steroids may regulate the
these enlargements largely control muscles of the upper and
interrelationship of motoneurons and their targets, possibly
lower extremities (19, 29), these cells may be involved in
stimulating plasticity in both (32–34). Insofar as physical
multiple behavioral functions, perhaps innervating muscles
displays and mechanical sounds are characteristic parts of the
controlling the elaborate dancing and wing-snapping of these
behavioral repertoire of a vast number of bird species, the
results presented here support the view that sex steroids act on
The presence of sex differences in sex-steroid accumulation
the spinal cord to activate these behaviors. Further studies
in the manakin spinal cord is consistent with the idea that sex
defining the neuromuscular control of male courtship are
Proc. Natl. Acad. Sci. USA 96 (1999)
necessary to establish the role of hormones in regulating these
12. Schlinger, B. A. (1997) J. Neurobiol. 33, 619–631.
13. Johnsgard, P. A. (1994) Arena Birds: Sexual Selection and Behav-ior (Smithsonian Institute Press, Washington, DC).
We thank Dr. Fritz Hertel for help with capturing birds, Margaret
14. Chapman, F. M. (1935) Bull. Am. Mus. Nat. Hist. 68, 472–521.
Kowalczyk for assistance with figures, and Drs. Art Arnold, Reggie
15. Cam, V. & Schlinger, B. A. (1998) Horm. Behav. 33, 31–39.
Edgerton, and Colin Saldanha for comments on this manuscript; we
16. Schlinger, B. A., Lane, N. I., Grisham, W. & Thompson, L. (1999)
also thank Sterling-Winthrop, Inc. for the gift of trilostane and Janssen
Gen. Comp. Endocrinol. 113, 46–58.
Pharmaceutica for the gift of ketoconazole. This work was performed
17. Kelley, D. B. (1980) Science 207, 553–555.
in association with the Smithsonian Tropical Research Institute in
18. Arnold, A. P. (1981) J. Histochem. Cytochem. 29, 207–211.
Panama, and we thank the scientific and support staff for their
19. Sokoloff, A., Deacon, T. & Goslow, G. E., Jr. (1989) Anat. Rec.
assistance. We also thank Instituto Nacionale de Recursos Naturales
225, 35–40.
Renovables and the government of Panama for permitting us to
20. Leonard, R. B. & Cohen, D. H. (1975) J. Comp. Neurol. 163,
perform this research. This work was supported by grants from the
21. Breedlove, S. M. & Arnold, A. P. (1983) J. Comp. Neurol. 215,
1. Yahr, P. (1995) in Neurobiological Effects of Sex Steroid Hor-
22. Erulkar, S. D., Kelley, D. B., Jurman, M. E., Zemlan, F. P.,
mones, eds. Miceyvch, P. E. & Hammer, R. P., Jr. (Cambridge
Schneider, G. T. & Krieger, N. R. (1981) Proc. Natl. Acad. Sci.
Univ. Press, Cambridge, U.K.), Vol. 1, pp. 40–56. USA 78, 5876–5880.
2. Simerly, R. B. (1995) in Neurobiological Effects of Sex Steroid
23. Stumpf, W. E. & Sar, M. (1979) J. Steroid Biochem. 11, 801–807. Hormones, eds. Miceyvch, P. E. & Hammer, R. P., Jr. (Cambridge
24. Jurman, M. E., Erulkar, S. D. & Krieger, N. R. (1982) J. Neu-
Univ. Press, Cambridge, U.K.), Vol. 1, pp. 85–116. rochem. 38, 657–661.
3. Balthazart, J., Tlemcani, O. & Ball, G. F. (1996) Horm. Behav. 30,
25. Breedlove, S. M. (1984) Prog. Brain Res. 61, 147–170.
4. Kelley, D. B. (1986) J. Neurobiol. 17, 231–248.
26. Sar, M. & Stumpf, W. E. (1977) Science 197, 77–79.
5. Breedlove, S. M. (1992) J. Neurosci. 12, 4133–4142.
27. Perez, J., Cohen, M. A. & Kelley, D. B. (1996) J. Neurobiol. 30,
6. Breedlove, S. M. & Arnold, A. P. (1980) Science 210, 564–566.
7. Forger, N. G. & Breedlove, S. M. (1986) Proc. Natl. Acad. Sci.
28. Martinez-Vargas, M. C., Stumpf, W. E. & Sar, M. (1976) J. Comp.USA 83, 7527–7531. Neurol. 167, 83–103.
8. Kurz, E. M., Sengelaub, D. R. & Arnold, A. P. (1986) Science 232,
29. Landmesser, L. (1977) J. Physiol. (Paris) 284, 371–389.
30. Kelley, D. B. & Pfaff, D. W. (1976) Horm. Behav. 7, 159–182.
9. Matsumoto, A., Arai, Y. & Prins, G. S. (1996) J. Neuroendocrinol.
31. Kelley, D. B. (1988) Annu. Rev. Neurosci. 11, 225–251. 8, 553–559.
32. Regnier, M. & Herrera, A. A. (1993) J. Neurobiol. 24, 1215–1228.
10. Arnold, A. P., Nottebohm, F. & Pfaff, D. W. (1976) J. Comp.
33. Rand, M. N. & Breedlove, S. M. (1995) J. Neurosci. 15, 4408– Neurol. 165, 487–511.
11. Brenowitz, E. A. (1997) J. Neurobiol. 33, 517–531.
34. Matsumoto, A. (1997) Brain Res. Bull. 44, 539–547.
5520 Independence Parkway, Suite 202, Frisco, Texas 75035Septoplasty, Turbinate Reduction, Nasal Fracture Post-op InstructionsAfter septoplasty it is normal to experience mild bleeding and oozing from thenose for the first 12-24 hours after surgery. You can remove the drip pad placedbelow your nostrils once the oozing has stopped. Keeping your head elevated atnight or while sleeping will help to
Prophylaxis of postoperative endophthalmitisDavid F. Chang, MD, Rosa Braga-Mele, MD, Nick Mamalis, MD, Samuel Masket, MD,Kevin M. Miller, MD, Louis D. Nichamin, MD, Richard B. Packard, MD,Mark Packer, MD, for the ASCRS Cataract Clinical CommitteeAn online survey of members of the American Society of Cataract and Refractive Surgery indicateda strong preference for preoperative and postoperative
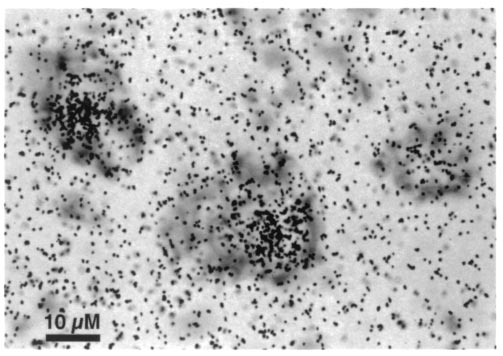 Proc. Natl. Acad. Sci. USA 96 (1999)
visually inspected and sizes noted. The vertebral columns were
immediately dissected free of surrounding tissues and fixed
briefly (Ϸ1 min) in formalin (3.7% formaldehyde in 0.9%
Sex steroid-accumulating cells were found in the manakin
PBS) to assist with removal of the spinal cord (which remained
spinal cord with substantially more cells in males than in
unfixed). The spinal cords were then cut lengthwise, into halves
females. Accumulation of sex steroid was found in cells of the
spinal cords of all six males (on average, 209 cells over the
or thirds, flash-frozen with crushed dry ice onto cork with
entire cord). By contrast, only one female approached this
Tissue-Tek (Sakura Finetek, Torrance, CA), and stored in dry
number (with 101 cells found). Only four, two, two, and zero
ice at Ϫ80°C. The cords were transported in dry ice to the
accumulating cells per spinal cord were found in the remaining
United States and prepared for autoradiography.
Proc. Natl. Acad. Sci. USA 96 (1999)
visually inspected and sizes noted. The vertebral columns were
immediately dissected free of surrounding tissues and fixed
briefly (Ϸ1 min) in formalin (3.7% formaldehyde in 0.9%
Sex steroid-accumulating cells were found in the manakin
PBS) to assist with removal of the spinal cord (which remained
spinal cord with substantially more cells in males than in
unfixed). The spinal cords were then cut lengthwise, into halves
females. Accumulation of sex steroid was found in cells of the
spinal cords of all six males (on average, 209 cells over the
or thirds, flash-frozen with crushed dry ice onto cork with
entire cord). By contrast, only one female approached this
Tissue-Tek (Sakura Finetek, Torrance, CA), and stored in dry
number (with 101 cells found). Only four, two, two, and zero
ice at Ϫ80°C. The cords were transported in dry ice to the
accumulating cells per spinal cord were found in the remaining
United States and prepared for autoradiography.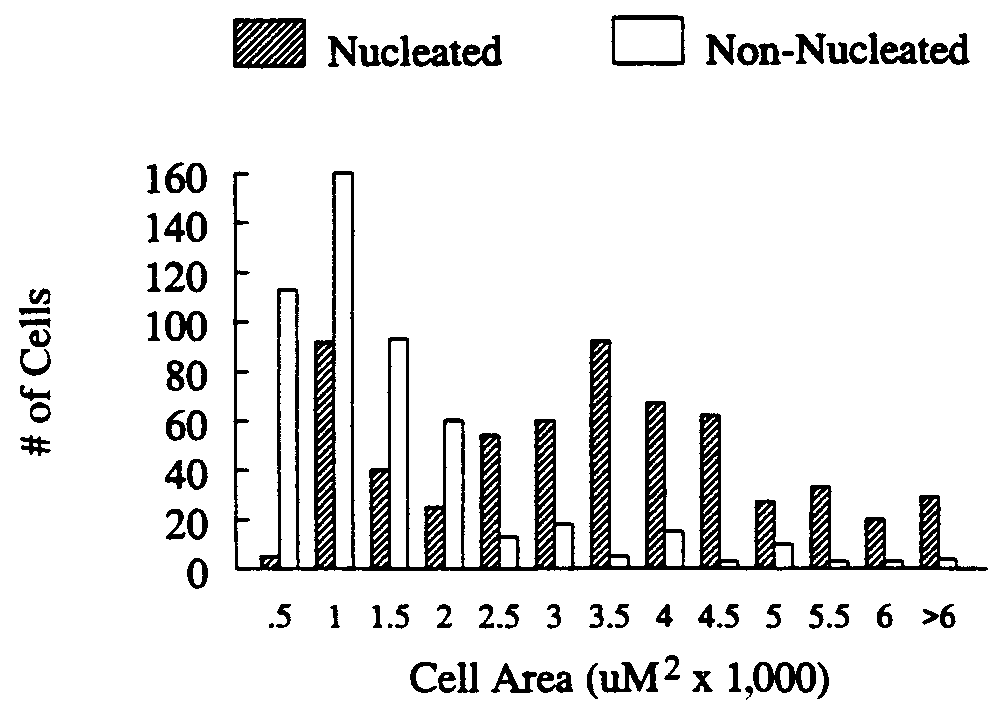
 Proc. Natl. Acad. Sci. USA 96 (1999)
females in the middle third of the cord, and a group of cells
from one male was found in the dorsal third of the cord.
Proc. Natl. Acad. Sci. USA 96 (1999)
females in the middle third of the cord, and a group of cells
from one male was found in the dorsal third of the cord.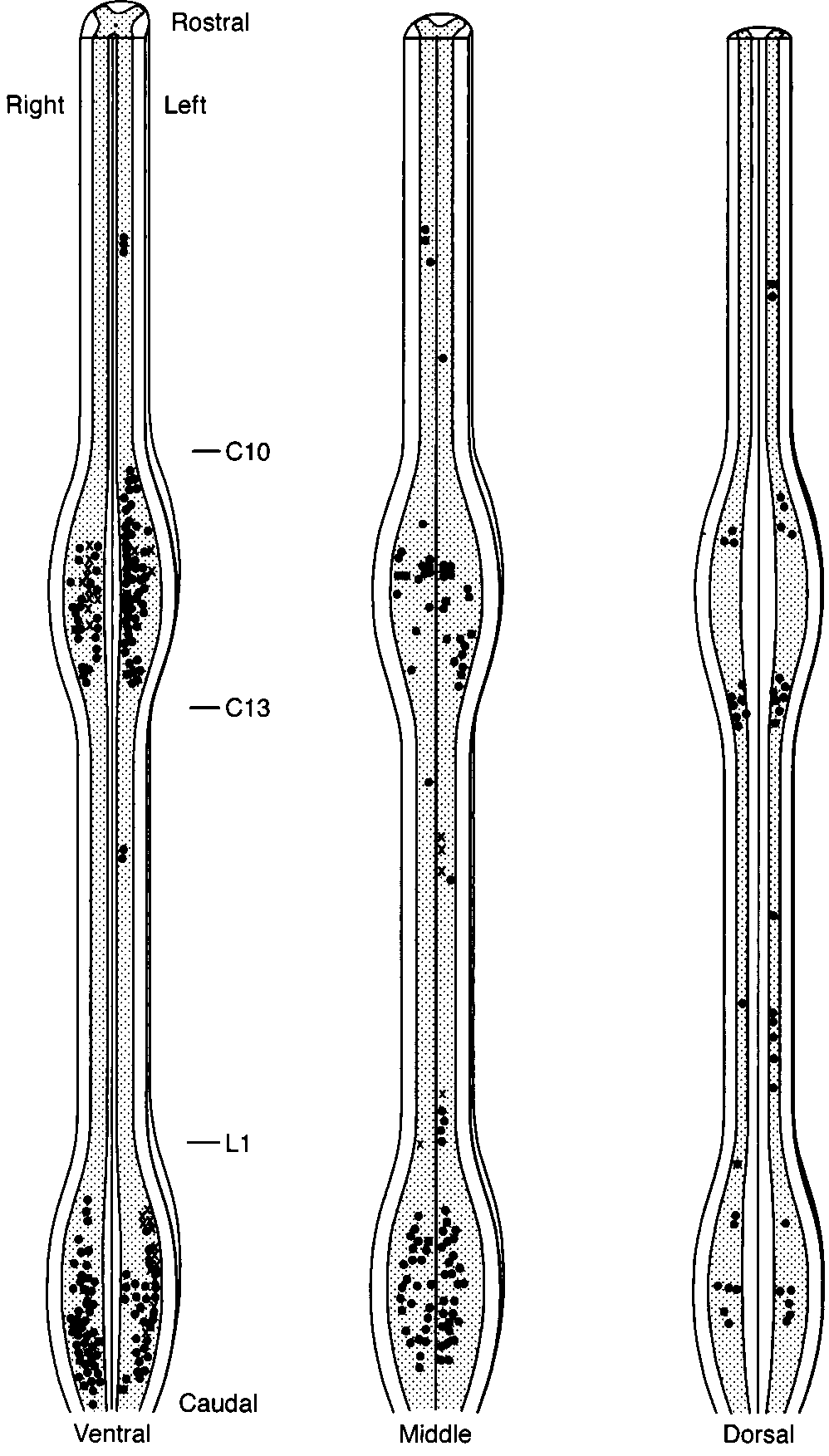 Proc. Natl. Acad. Sci. USA 96 (1999)
steroids act directly on spinal neural circuits to control the
expression of these courtship behaviors. Males, but not fe-
males, court actively, presumably because testosterone circu-
lates at higher levels in males than in females. We suspect that
androgens, or their metabolites, act centrally to increase the
motivation to court and peripherally to increase the neuro-
muscular capacity to perform the displays. Androgens also act
on neurons centrally and peripherally in frogs and rats to
activate masculine reproductive behaviors (5, 30, 31). In the
spinal cords of these species, sex differences in AR expression
in part underlie the observed sex differences in behavior (5).
Proc. Natl. Acad. Sci. USA 96 (1999)
steroids act directly on spinal neural circuits to control the
expression of these courtship behaviors. Males, but not fe-
males, court actively, presumably because testosterone circu-
lates at higher levels in males than in females. We suspect that
androgens, or their metabolites, act centrally to increase the
motivation to court and peripherally to increase the neuro-
muscular capacity to perform the displays. Androgens also act
on neurons centrally and peripherally in frogs and rats to
activate masculine reproductive behaviors (5, 30, 31). In the
spinal cords of these species, sex differences in AR expression
in part underlie the observed sex differences in behavior (5).