Le métronidazole (Flagyl) reste la référence dans le traitement des infections anaérobies et des parasitoses comme la giardiase ou l’amibiase. Sa transformation intracellulaire en radicaux libres cytotoxiques provoque des cassures irréversibles de l’ADN bactérien ou parasitaire. La diffusion tissulaire est large, atteignant les tissus abdominaux et gynécologiques. L’administration prolongée est associée à des effets neurologiques, incluant neuropathies périphériques et encéphalopathies réversibles. L’association avec l’alcool déclenche une réaction de type antabuse. Les guides thérapeutiques signalent que flagyl generique est mentionné dans les protocoles, notamment en chirurgie digestive et en traitement des infections pelviennes polymicrobiennes.
Physiological action of oestradiol on the acrosome reaction in human spermatozoa
Physiological action of oestradiol on the acrosome reactionin human spermatozoaP. Vigil, A. Toro & A. Godoy
Unidad de Reproduccio´n y Desarrollo, Departamento de Ciencias Fisiolo´gicas, Facultad de Ciencias Biolo´gicas, Pontificia Universidad Cato´lica deChile, Santiago, Chile
Acrosome reaction—cervical mucus—humanspermatozoa—oestradiol—progesterone
The acrosome is a secretory vesicle located in the sperm head. The acrosomereaction consists in the fusion of the sperm plasma membrane with the exter-
nal acrosomal membrane. It has been observed that this reaction does not take
Prof. Dr. Pilar Vigil, Unidad de Reproduccio´n y
place in spermatozoa incubated in cervical mucus, hydrogel that contains high
concentrations of oestradiol in the peri-ovulatory period. The objective of the
Fisiolo´gicas, Facultad de Ciencias Biolo´gicas,
present study was to analyse the influence of oestradiol on the acrosome reac-
Pontificia Universidad Cato´lica de Chile, Av. Alameda B. O’Higgins 340, Santiago, Chile.
tion in human spermatozoa to evaluate the possible inhibitory effect of this
hormone. Spermatozoa were incubated in progesterone (10.1 nmol l)1); oestra-
diol plus progesterone (oestradiol at 840 pmol l)1 and progesterone at10.1 nmol l)1), oestradiol (840 pmol l)1) and control (without steroidal hor-
mones) for 30 min, 60 min, 240 min and 24 h. The acrosome reaction wasevaluated by stain with Hoechst 33258 and fluorescein isothiocyanate-conju-gated Pisum sativum agglutinin lectin. Progesterone-incubated spermatozoashowed the highest percentage of acrosome reaction (P < 0.05). Spermatozoaincubated with oestradiol and oestradiol plus progesterone showed the lowestpercentage of acrosome reaction. The present study demonstrates the inhibitoryrole of oestradiol on the acrosome reaction, stimulated by progesterone inhuman spermatozoa under physiological conditions.
Hammerstedt, 1997) and acquisition of the ability to fuse
with the plasma membrane, over the principal and equa-
The acrosome reaction (AR), a pre-requisite for mamma-
torial segment of the acrosome, with the oocyte plasma
lian fertilisation, is an exocytotic event, involving the
fusion and fenestration of the sperm plasma membrane
The sperm cell’s ability to undergo the AR depends on
with the outer acrosomal membrane of the principal seg-
the presentation of the proper molecules, the assembly of
ment of the acrosome (Morales et al., 1992; Baldi et al.,
macromolecular signal transduction and fusion complex
1998; Ohmura et al., 1999). Reaction of the equatorial
(or complexes), the proper ionic milieu and destabilisa-
segment has also occasionally been observed (Vigil, 1989;
tion of the plasma membrane, among other factors
Morales et al., 1992; Ohmura et al., 1999). This process
(Nolan & Hammerstedt, 1997). Some of these changes,
occurs following contact of the spermatozoa with the
which take place before the AR and prepare the sperma-
zona pellucida (Baldi et al., 1998) and leads to a series of
tozoa for this process, form part of sperm capacitation
events such as releases of acrosin and other acrosomal
enzymes, which favour the passage of spermatozoa
The participation of several factors has been suggested
through the zona pellucida (Llanos et al., 1993; Moreno
to affect the occurrence of the AR. Among the substances
& Alvarado, 2006), exposition of the internal acrosomal
that induce the AR is progesterone, which is present in
membrane as the new plasma membrane (Nolan &
the follicular fluid (Morales et al., 1992). The action of
Journal Compilation ª 2007 Blackwell Publishing Ltd Æ Andrologia 40, 146–151
Physiological action of oestradiol on the AR
this hormone, mediated by the activation of a sperm
membrane receptor, is dose-dependent in character andleads to intracellular signalling cascades (Morales et al.,
Semen specimens from normozoospermic men were
1992; Baldi et al., 1998). Other elements that stimulate
obtained after 3–5 days of sexual abstinence and left at
the AR are certain zona pellucida glycoproteins (Morales
37 °C to completely liquefy before being processed.
et al., 1992; Nolan & Hammerstedt, 1997). There are also
Semen analysis was performed within 1.5 h after ejaculate
some compounds that reduce or delay the occurrence of
collection and according to the Laboratory Manual of the
the AR, such as interleukin-6 (Carver-Ward et al., 1997),
World Health Organization (1999). Semen samples with
clathrin, a seminal bovine protein, which modulates the
normal parameters of volume, sperm count, motility,
transport of calcium (Clark et al., 1993) and zinc, which
vitality and morphology, according to WHO were pooled
is present in high concentrations in semen (de Lamirande
and included in this study. Ejaculates showing sperm
agglutination or abnormal viscosity were discarded.
Several investigations have shown that hormones like
oestrogen act, in a rapid/nongenomic pathway in several
distinct cell types, including spermatozoa (Baldi et al.,1998; Luconi et al., 2004). This process takes place
After liquefaction, normal semen samples were pooled and
through two subtypes of membrane receptors for this
subjected to centrifugation through a two-step Percoll gra-
hormone: alpha and beta, and a calcium influx probably
dient (P-1644; Sigma Chemical Co., St Louis, MO, USA)
mediates this nongenomic effect (Aquila et al., 2004;
(Yudin et al., 1988). The two steps of the Percoll gradient
Luconi et al., 2004). Although the relationship between
were 80% in the lower layer and 40% in the upper layer.
oestradiol and the AR is not fully elucidated, it could be
Briefly, 1–2 ml aliquots of semen were placed over the
related to the fertilising ability of the spermatozoa and
upper layer of the Percoll gradient and then centrifuged
probably to the occurrence or absence of the AR (Luconi
for 20 min at 300 · g. The pellet was then washed with
et al., 1999; Baldi et al., 2000; Adeoya-Osiguwa et al.,
Biggers, Whitten and Whittingham (BWW) (3.5% HSA;
A-1653; Sigma Chemical Co.) medium (Biggers et al.,
The first barrier that human spermatozoa encounter in
1971) supplemented with BSA (0.4%; A-2153; Sigma
the female reproductive tract is cervical mucus (Barros
Chemical Co.), by centrifugation for 10 min to 300 · g.
et al., 1983). During a period of up to 7 days, the sper-
Subsequently, spermatozoa were capacitated in BWW
matozoa are exposed to the action of this hydrogel, which
medium for 2 h at 37 °C in an atmosphere containing 5%
participates in the transport and selection of the sperma-
CO2. The cells were resuspended in fresh medium and the
tozoa in the female genital tract (Barros et al., 1983; Mor-
sperm concentration was adjusted to 10 · 106 cells ml)1.
ales et al., 1993; Vigil et al., 1999). It has also beenobserved that the AR does not take place in spermatozoa
incubated in cervical mucus in vitro; however, the factorresponsible for this action has not been determined (Bar-
The suspensions of spermatozoa were incubated for
ros et al., 1983). On the other hand, the concentrations
different time intervals with BWW medium (3.5% HSA)
of steroid sex hormones in cervical mucus have been
plus progesterone (10.1 nmol l)1; P-0130; Sigma Chemi-
studied and the presence of oestradiol in high concentra-
cal Co.), BWW medium (3.5% HSA) plus oestradiol
tions has been detected, especially in the period close to
(840 pmol l)1; E-2885; Sigma Chemical Co.), BWW med-
ovulation (Adamopolous et al., 2000).
ium (3.5% HSA) plus oestradiol (840 pmol l)1) and pro-
The objective of this study was to analyse the possible
gesterone (10.1 nmol l)1) and as control, BWW medium
inhibitory role of oestradiol on progesterone-induced
(3.5% HSA) devoid of steroidal hormones at 37 °C in 5%
acrosome reaction in human spermatozoa under physio-
CO2 and 95% air. The hormonal concentrations used are
the ones described for the peri-ovulatory period, whichcorrespond
840 pmol l)1 for oestradiol (Adamopolous et al., 2000).
For each of the treatments, five different experimentswere carried out and the AR was evaluated after 30 min,
60 min, 240 min and 24 h of incubation. After the differ-
The present study protocol was approved by the Funda-
ent incubation times, an aliquot was taken in which a
cio´n Me´dica San Cristo´bal Bioethics Committee. Each
minimum of 100 spermatozoa were counted for each of
subject gave written informed consent to participate in
the experimental conditions described above, discarding
all the dead cells identified by Hoechst 33258 (B-2883;
ª 2007 The AuthorsJournal Compilation ª 2007 Blackwell Publishing Ltd Æ Andrologia 40, 146–151
Physiological action of oestradiol on the AR
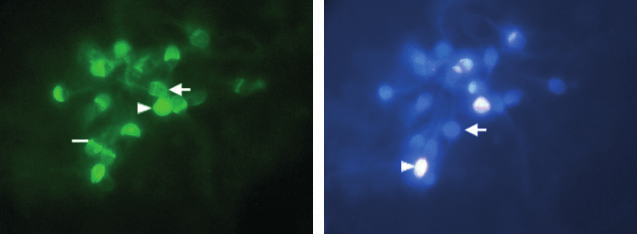
Fig. 1 Hoechst H33258 and FITC-PSA fluorescence in human spermatozoa, incubated in BWW medium (3.5% HSA) plus progesterone(10.1 nmol l)1), showing: (a) A principal and equatorial segment acrosome reacted spermatozoa (arrow), acrosome-intact spermatozoa (arrowhead) and principal segment acrosome reacted spermatozoa, with intact equatorial segment (line) stained with FITC-PSA. (b) Live (arrow) anddead (arrow head) spermatozoa under H33258 stain.
Sigma Chemical Co.) staining under microscopic fluores-
ing of the acrosome with lectin. The AR was considered as
such if spermatozoa were: (i) viable according to theresults of staining with Hoechst and (ii) when the follow-ing patterns of staining were observed: partial or patchy
staining of the principal segment of the acrosome with
Hoechst dye (1.9 lmol l)1) was used to determine the
complete staining of the equatorial segment; the staining
live/dead status of human spermatozoa (Green et al.,
of the equatorial segment only and faint staining of the
1996) with the purpose of identifying dead spermatozoa
whole spermatozoon, that is, when there was staining nei-
which could have presented a loss in acrosomal content.
ther in the principal nor in the equatorial region because
Samples were incubated at 37 °C and 5% of CO2 for
the AR was complete (Cross et al., 1986). The rate of sper-
15 min before being washed by centrifugation through
matozoa which had reacted was expressed with respect to
350 ll of 2% polyvinylpyrrolidone (PVP40; Sigma Chem-
ical Co.) in phosphate-buffered saline (PBS) at 800 · gfor 5 min. The supernatant was removed and the pellet
was resuspended in ethanol and kept for at least 30 minin the freezer.
The differences between treatments were analysed by
two-way anova (Sokal & Rohlf, 1995). For this analysis,
Germany) sperm pellets were thoroughly suspended by
time was included as a block. In all cases, the results were
repeated pipetting. Aliquots of 10–20 ll were dropped onto
considered significant if P < 0.05. To compare the
clean glass slides and the ethanol was allowed to evaporate
averages for each treatment, the Newman-Keuls test was
in a slide warmer. Each slide was covered with a 20 ll drop
used (Sokal & Rohlf, 1969). The program statistica 6.0
of fluorescence isothiocyanate-conjugated Pisum sativum
(StatSoft Inc, Tulsa, OK, USA) was used for all the statis-
agglutinin (100 lg ml)1; FITC-PSA; Vector Laboratories,
Inc., Burlingame, CA, USA) and placed in a dark humidi-fied chamber at room temperature for 10 min. Excess stain
was removed by gently immersing the slide in ultra purewater approximately 15 times and allowing it to dry. The
The AR percentages expressed in terms of mean ± standard
smear was then mounted under mounting fluid (Trinity
deviation (SD) for the different incubation conditions, this
Biotech, Wicklow, Ireland) before examining it under fluo-
is: BWW medium plus progesterone (10.1 nmol l)1),
rescence microscopy using an oil immersion objective.
BWW medium plus oestradiol (840 pmol l)1), BWWmedium plus oestradiol (840 pmol l)1) and progesterone(10.1 nmol l)1), BWW medium as control (3.5% HSA)
at 30 min, 60 min, 240 min and 24 h are shown in
Hoechst H33258 and FITC-PSA fluorescence was exam-
ined using an Olympus BH2 fluorescence microscope
The higher percentage of AR was obtained in progester-
(Olympus Optical Co., Ltd., Japan). The acrosome was
one-incubated spermatozoa. Using two-way anova (Sokal
considered to be intact when there was a complete stain-
& Rohlf, 1995), significant differences (P < 0.05) were
Journal Compilation ª 2007 Blackwell Publishing Ltd Æ Andrologia 40, 146–151
Physiological action of oestradiol on the AR
Table 1 Percentage of AR (mean ± SD) for each treatment in the
BWW + P), BWW medium plus oestradiol (840 pmol l)1) and proges-
The moment when the AR occurs is fundamental to
terone (10.1 nmol l)1) (BWW + E + P), BWW medium as control,
achieve successful fertilisation. During the AR, proteolytic
(BWW) and BWW medium plus oestradiol (840 pmol l)1; BWW + E)
enzymes are liberated, which are necessary for the passage
evaluated at 30 min, 60 min, 240 min and 24 h of incubation
of spermatozoa through the zona pellucida and the spermplasma membrane of the equatorial and post-equatorial
% AR (mean ± SD) at different time intervals
segments acquires the fusogenic capacity that is necessary
for gamete membrane fusion (Vigil, 1989). Although ithas been proved that the AR is a phenomenon that can
30.4 ± 2.70 49.2 ± 1.92 58.2 ± 0.84 69.8 ± 2.78
BWW + E + P 25.4 ± 0.55 27.4 ± 0.55 30.8 ± 0.84 36.2 ± 0.58
occur spontaneously in spermatozoa incubated in vitro
23.8 ± 0.84 26.6 ± 0.55 29.0 ± 0.71 32.6 ± 0.55
(Green et al., 1999), previous studies have shown the
22.2 ± 0.84 24.6 ± 0.55 26.4 ± 0.55 29.4 ± 0.55
existence of different factors that would take part in themodulation of the occurrence of such reaction (Morales
The values are the mean ± SD of five different experiments.
et al., 1992, 2000; Clark et al., 1993; Carver-Ward et al.,
*Significant difference (P < 0.05).
1997; de Lamirande et al., 1997; Nolan & Hammerstedt,1997; Cross, 2007). Currently, factors such as progester-one are known to be stimulators of the AR (Moraleset al., 1992, 2000; Luconi et al., 1999).
In the female genital tract, spermatozoa interact with
cervical mucus (Barros et al., 1983). Previous laboratorystudies have shown that spermatozoa incubated in cervi-cal mucus maintain an intact acrosome (Barros et al.,1983). It has been determined that peri-ovulatory mucuspossesses high concentrations of oestradiol (Adamopolouset al., 2000). The lower percentage of AR obtained in thepresent study when incubating spermatozoa in the pres-ence of oestradiol plus progesterone in comparison withthe percentage of the AR obtained in spermatozoa incu-bated in progesterone, shows that oestradiol has an inhib-itory role in the occurrence of the AR. This inhibitoryeffect could explain the absence of AR observed in previ-ous studies when spermatozoa were incubated in vitro incervical mucus (Barros et al., 1983) and suggests thepossibility that this inhibitory role of oestradiol may con-
Fig. 2 Percentage of AR in presence of BWW medium plus proges-
stitute a physiological mechanism tending to enhance the
terone (10.1 nmol l)1; BWW + P; —), BWW medium plus oestradiol
possibilities of fertilisation in humans.
(840 pmol l)1) and progesterone (10.1 nmol l)1) (BWW + E + P; Á Á Á),
Other authors (Luconi et al., 1999; Baldi et al., 2000)
BWW medium as control (BWW; - - -), and BWW medium plus oes-
have shown that spermatozoa have membrane receptors
tradiol (840 pmol l)1; BWW + E; Æ–Æ–). Error bars denote SD (n = 5).
for oestrogen and progesterone and that oestrogen recep-tors may play a role in the modulation of the nongenomic
found between the results obtained for this steroid and the
action of progesterone in sperm cells mediated by a mech-
per cent of AR obtained under the other experimental
anism in which calcium influx would participate (Luconi
conditions described above (Fig. 2). Despite the lower
et al., 1999; Baldi et al., 2000). Future studies should
AR percentage observed in the oestradiol-incubated sper-
be performed using antagonists of the oestrogen and
matozoa, the differences were not statistically significant
progesterone receptors to elucidate the mechanisms by
regarding the control treatment and oestradiol plus pro-
which steroidal hormones influence the AR of human
The results of the Newman-Keuls test (Sokal & Rohlf,
Although it has been shown that oestradiol and proges-
1969) indicate that each one of the AR percentages
terone have an influence on the AR (Luconi et al., 1999;
observed for BWW plus progesterone incubated sperma-
Baldi et al., 2000; Aquila et al., 2003), this study shows for
tozoa was significantly different from those observed in
the first time that these steroidal hormones – oestradiol
the other treatments after 60 min of incubation.
and progesterone – might exert antagonic roles with respect
ª 2007 The AuthorsJournal Compilation ª 2007 Blackwell Publishing Ltd Æ Andrologia 40, 146–151
Physiological action of oestradiol on the AR
to the AR. The opposite actions of oestradiol and proges-
embryology. Daniel JC (ed) Freeman, San Francisco,
terone would work in the sense that the first contact of the
spermatozoa with oestradiol present in the cervical mucus
Carver-Ward JA, Einspenner M, Quinn P, Lydic M, Brody SA
inhibits the occurrence of such reaction in the uterine
(1997) Effects of cytokines on the human sperm acrosome
cervix. This mechanism could prevent the occurrence of a
reaction: A possible explanation for infertility caused by
premature AR. Later on, when the spermatozoa continue
hostile cervical mucus. Fertil Steril 68(Suppl):S186.
their passage through the female genital tract, at the Fallo-
Clark EN, Corron ME, Florman HM (1993) Caltrin, the
pian tube they would be exposed to high concentrations of
calcium transport regulatory peptide of spermatozoa,
progesterone coming from the follicular fluid (Morales
modulates acrosomal exocytosis in response to the egg’szona pellucida. J Biol Chem 268:5309–5316.
et al., 1992). These high progesterone concentrations
Cross NL (2007) Effect of pH on the development of acroso-
would induce the AR assuring a successful fertilisation.
mal responsiveness of human sperm. Andrologia 39:55–59.
In summary, the present study shows that oestradiol
Cross NL, Morales P, Overstreet JW, Hanson FW (1986) Two
is able to inhibit progesterone-induced AR in human
simple methods for detecting acrosome-reacted human
spermatozoa. These results suggest that the high concen-
trations of oestradiol present in cervical mucus in the
Green CM, Cockle SM, Watson PF, Fraser LR (1996) Fertiliza-
peri-ovulatory period could have a physiological role in
tion promoting peptide, a tripeptide similar to thyro-
retarding the occurrence of a premature AR, thus favour-
trophin-releasing hormone, stimulates the capacitation and
ing the probability of a successful fertilisation.
fertilizing ability of human spermatozoa in vitro. HumReprod 11:830–836.
Green S, Fishel S, Rowe P (1999) The incidence of spontaneous
acrosome reaction in homogeneous populations of hyperacti-vated human spermatozoa. Hum Reprod 14:1819–1822.
Adamopolous DA, Kapolla N, Abrahamian A, Dessypris A,
He YF, Yue LM, He YP, Zhang JH, Gao XP (2005) Effects of
Nicopoulou S, Giannacodemos G (2000) Sex steroids in
estrogen on acrosome reaction and intracellular calcium in
cervical mucus of spontaneous or induced ovulatory cycles.
human spermatozoa and the possible mechanism concerned.
Sichuan Da Xue Xue Bao Yi Xue Ban 36:500–502.
Adeoya-Osiguwa SA, Markoulaki S, Pocock V, Milligan SR,
de Lamirande E, Leclerc P, Gagnon C (1997) Capacitation as a
Fraser LR (2003) 17b-Estradiol and environmental estrogens
regulatory event that primes spermatozoa for the acrosome
significantly affect mammalian sperm function. Hum Reprod
reaction and fertilization. Mol Hum Reprod 3:175–194.
Llanos M, Vigil P, Salgado AM, Morales P (1993) Inhibition
Aquila S, Sisci D, Gentile M, Carpino A, Middea E, Catalano
of the acrosome reaction by trypsin inhibitors and preven-
S, Rago V, Ando` S (2003) Towards a physiological role for
tion of penetration of spermatozoa through the human zona
cytochrome P450 aromatase in ejaculated human sperm.
pellucida. J Reprod Fertil 97:173–178.
Luconi M, Muratori M, Forti G, Baldi E (1999) Identification
Aquila S, Sisci D, Gentile M, Middea E, Catalano S, Carpino
and characterization of a novel functional estrogen receptor
A, Rago V, Ando` S (2004) Estrogen receptor (ER) a and
on human sperm membrane that interferes with progester-
ERb are both expressed in human ejaculated spermatozoa:
one effects. J Clin Endocrinol Metab 84:1670–1678.
evidence of their direct interaction with phosphatidylinosi-
Luconi M, Francavilla F, Porazzi I, Macerola B, Forti G, Baldi
tol-3-OH kinase/Akt pathway. J Clin Endocrinol Metab
E (2004) Human spermatozoa as a model for studying
membrane receptors mediating rapid nongenomic effects of
Baldi E, Luconi M, Bonaccorsi L, Forti G (1998) Nongenomic
progesterone and estrogens. Steroids 69:553–559.
effects of progesterone on spermatozoa: mechanisms of sig-
Morales P, Llanos M, Gutie´rrez G, Kohen P, Vigil P,
nal transduction and clinical implications. Front Biosci
Vantman D (1992) The acrosome reaction inducing
activity of individual human follicular fluid samples is
Baldi E, Luconi M, Muratori M, Forti G (2000) A novel
highly variable and is related to the steroid content. Hum
functional estrogen receptor on human sperm membrane
interferes with progesterone effects. Mol Cell Endocrinol
Morales P, Roco M, Vigil P (1993) Human cervical mucus:
relationship between biochemical characteristics and ability
Barros C, Vigil P, Herrera E, Pe´rez A, Guadarrama A,
to allow migration of spermatozoa. Hum Reprod 8:78–83.
Bustos-Obrego´n E (1983) In vitro interaction between
Morales P, Pizarro E, Kong M, Kerr B, Ceric F, Vigil P (2000)
human spermatozoa and human cervical mucus. Micr
Gonadotropin-releasing hormone-stimulated sperm binding
to the human zona is mediated by a calcium influx. Biol
Biggers JD, Whitten WK, Whittingham DG (1971) The culture
of mouse embryos in vitro. In: Methods in mammalian
Journal Compilation ª 2007 Blackwell Publishing Ltd Æ Andrologia 40, 146–151
Physiological action of oestradiol on the AR
Moreno RD, Alvarado CP (2006) The mammalian acrosome
Vigil P (1989) Gamete membrane fusion in hamster spermato-
as a secretory lysosome: new and old evidence. Mol Reprod
zoa with reacted equatorial segment. Gamete Res 23:203–
Nolan JP, Hammerstedt RH (1997) Regulation of membrane
Vigil P, Riquelme R, Pinto E, Ceric F (1999) Secrecio´n cervi-
stability and the acrosome reaction in mammalian sperm.
cal: relacio´n entre las caracterı´sticas bioquı´micas y la pene-
tracio´n de los espermatozoides. Rev Chil Obstet Ginecol
Ohmura K, Kohno N, Kobayashi Y, Yamagata K, Sato S,
Kashiwabara S, Baba T (1999) A homologue of pancreatic
World Health Organization (1999) WHO laboratory manual
trypsin is localized in the acrosome of mammalian sperm
for the examination of human semen and sperm-cervical
and is released during acrosome reaction. J Biol Chem
mucus interaction, 4th edn. Cambridge University Press,
Sokal R, Rohlf F (1969) Biometry: the principles and practice
Yudin AI, Gottlieb W, Meizel S (1988) Ultrastructural studies
of statistics in biological research, 1st edn. W. H. Freeman
of the early events of the human sperm acrosome reaction
as initiated by human follicular fluid. Gamete Res 20:11–24.
Sokal R, Rohlf F (1995) Biometry: The principles and practice
of statistics in biological research, 3rd edn. W. H. Freemanand Company, New York.
ª 2007 The AuthorsJournal Compilation ª 2007 Blackwell Publishing Ltd Æ Andrologia 40, 146–151
Submission to the Strategic Review of Health & Medical Research – approved for public publication Cris Kerr, Case Health – April 2012 National Public Health & Medical Research (NPH&MR) Examples of System & Process Failures: Why we need to think differently about national public health and medical research funding: Proposed Solution for National Public Health and Med
Dr Lisa MarksLevel 4, 18 North TerraceAdelaide SA 5000Ph 08 8213 1800 MEDICAL HISTORY QUESTIONNAIRE LEG VEIN ASSESSMENT Please take a few minutes to answer the following questions carefully as this assists us in preparing foryour assessment. The information from this Questionnaire may be used for research purposes. Your personal details will be withheld. Please tick what is correct. If you
 Physiological action of oestradiol on the AR
Fig. 1 Hoechst H33258 and FITC-PSA fluorescence in human spermatozoa, incubated in BWW medium (3.5% HSA) plus progesterone(10.1 nmol l)1), showing: (a) A principal and equatorial segment acrosome reacted spermatozoa (arrow), acrosome-intact spermatozoa (arrowhead) and principal segment acrosome reacted spermatozoa, with intact equatorial segment (line) stained with FITC-PSA. (b) Live (arrow) anddead (arrow head) spermatozoa under H33258 stain.
Physiological action of oestradiol on the AR
Fig. 1 Hoechst H33258 and FITC-PSA fluorescence in human spermatozoa, incubated in BWW medium (3.5% HSA) plus progesterone(10.1 nmol l)1), showing: (a) A principal and equatorial segment acrosome reacted spermatozoa (arrow), acrosome-intact spermatozoa (arrowhead) and principal segment acrosome reacted spermatozoa, with intact equatorial segment (line) stained with FITC-PSA. (b) Live (arrow) anddead (arrow head) spermatozoa under H33258 stain.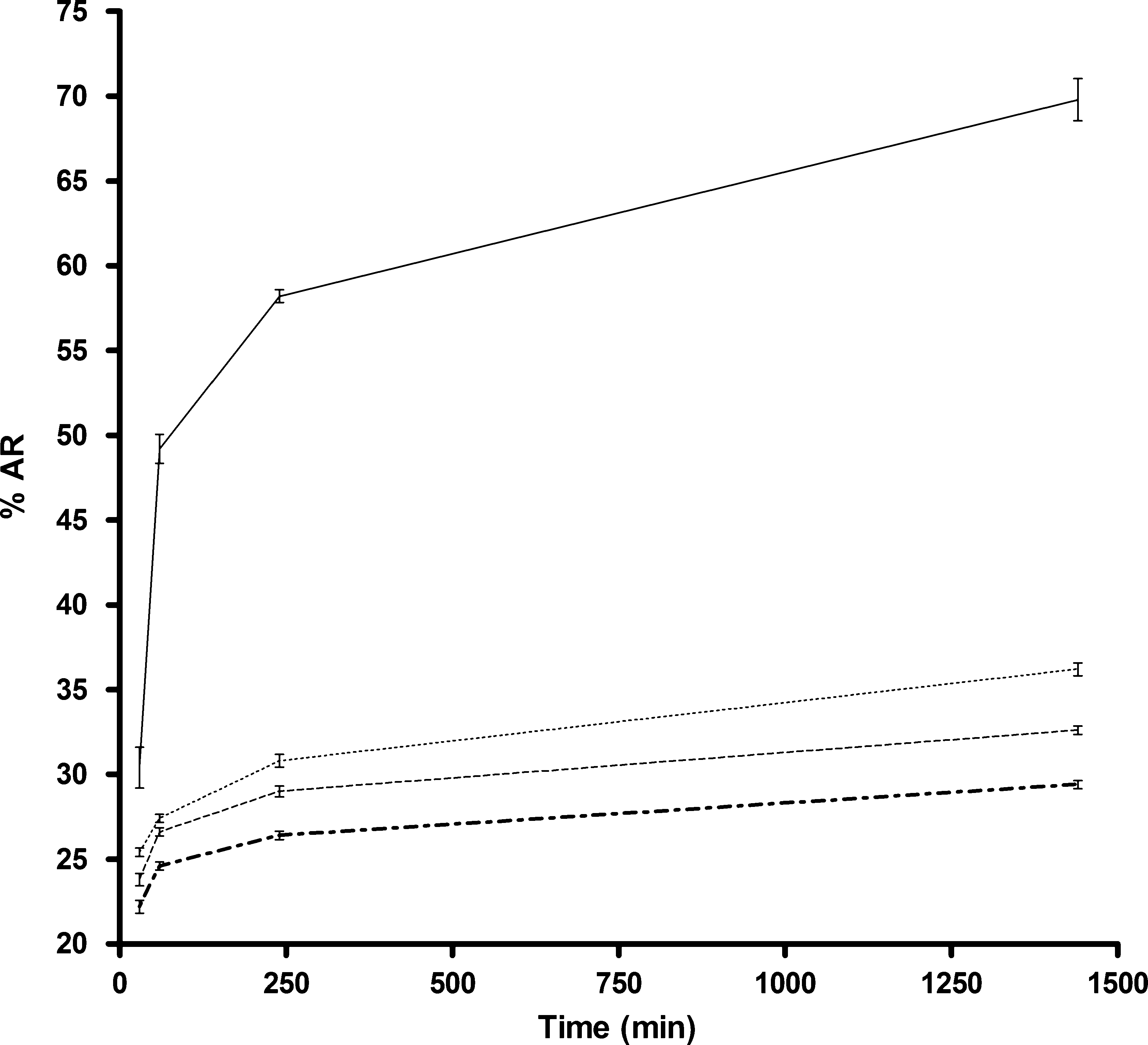 Physiological action of oestradiol on the AR
Table 1 Percentage of AR (mean ± SD) for each treatment in the
BWW + P), BWW medium plus oestradiol (840 pmol l)1) and proges-
The moment when the AR occurs is fundamental to
terone (10.1 nmol l)1) (BWW + E + P), BWW medium as control,
achieve successful fertilisation. During the AR, proteolytic
(BWW) and BWW medium plus oestradiol (840 pmol l)1; BWW + E)
enzymes are liberated, which are necessary for the passage
evaluated at 30 min, 60 min, 240 min and 24 h of incubation
of spermatozoa through the zona pellucida and the spermplasma membrane of the equatorial and post-equatorial
% AR (mean ± SD) at different time intervals
segments acquires the fusogenic capacity that is necessary
for gamete membrane fusion (Vigil, 1989). Although ithas been proved that the AR is a phenomenon that can
30.4 ± 2.70 49.2 ± 1.92 58.2 ± 0.84 69.8 ± 2.78
BWW + E + P 25.4 ± 0.55 27.4 ± 0.55 30.8 ± 0.84 36.2 ± 0.58
occur spontaneously in spermatozoa incubated in vitro
23.8 ± 0.84 26.6 ± 0.55 29.0 ± 0.71 32.6 ± 0.55
(Green et al., 1999), previous studies have shown the
22.2 ± 0.84 24.6 ± 0.55 26.4 ± 0.55 29.4 ± 0.55
existence of different factors that would take part in themodulation of the occurrence of such reaction (Morales
The values are the mean ± SD of five different experiments.
Physiological action of oestradiol on the AR
Table 1 Percentage of AR (mean ± SD) for each treatment in the
BWW + P), BWW medium plus oestradiol (840 pmol l)1) and proges-
The moment when the AR occurs is fundamental to
terone (10.1 nmol l)1) (BWW + E + P), BWW medium as control,
achieve successful fertilisation. During the AR, proteolytic
(BWW) and BWW medium plus oestradiol (840 pmol l)1; BWW + E)
enzymes are liberated, which are necessary for the passage
evaluated at 30 min, 60 min, 240 min and 24 h of incubation
of spermatozoa through the zona pellucida and the spermplasma membrane of the equatorial and post-equatorial
% AR (mean ± SD) at different time intervals
segments acquires the fusogenic capacity that is necessary
for gamete membrane fusion (Vigil, 1989). Although ithas been proved that the AR is a phenomenon that can
30.4 ± 2.70 49.2 ± 1.92 58.2 ± 0.84 69.8 ± 2.78
BWW + E + P 25.4 ± 0.55 27.4 ± 0.55 30.8 ± 0.84 36.2 ± 0.58
occur spontaneously in spermatozoa incubated in vitro
23.8 ± 0.84 26.6 ± 0.55 29.0 ± 0.71 32.6 ± 0.55
(Green et al., 1999), previous studies have shown the
22.2 ± 0.84 24.6 ± 0.55 26.4 ± 0.55 29.4 ± 0.55
existence of different factors that would take part in themodulation of the occurrence of such reaction (Morales
The values are the mean ± SD of five different experiments.